



版权说明:本文档由用户提供并上传,收益归属内容提供方,若内容存在侵权,请进行举报或认领
文档简介
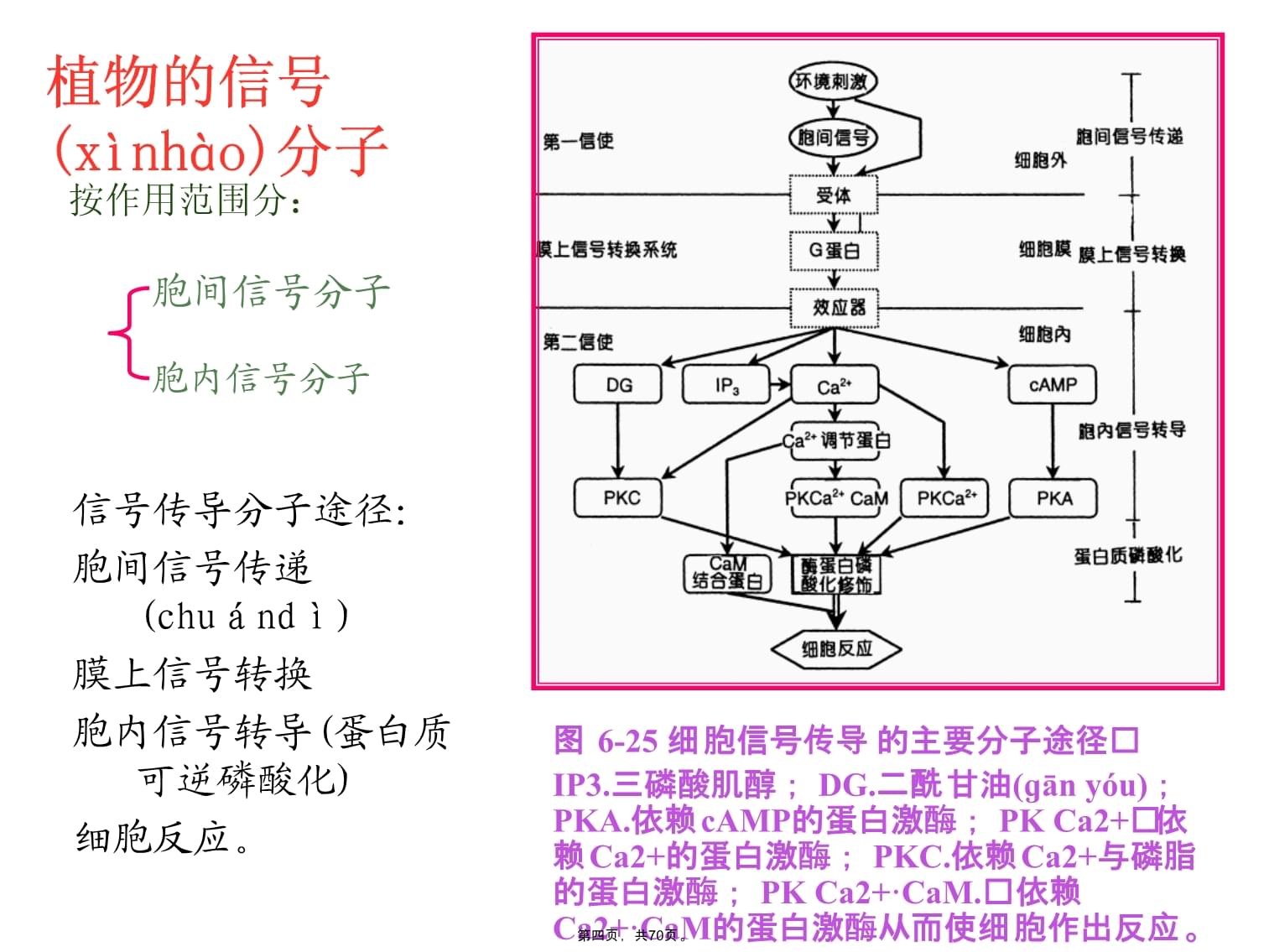
植物(zhíwù)细胞信号转导第一页,共70页。第一节植物体内(tǐnèi)的信号传导生长发育是基因在一定时间、空间上顺序表达的过程(guòchéng),而基因表达除受遗传信息支配外,还受环境的调控。植物在整个生长发育过程(guòchéng)中,受到各种内外因素的影响,这就需要植物体正确地辨别各种信息并作出相应的反应,以确保正常的生长和发育。例如植物的向光性能促使植物向光线充足的方向生长,在这个过程(guòchéng)中,首先植物体要能感受到光线,然后把相关的信息传递到有关的靶细胞,并诱发胞内信号转导,调节基因的表达或改变酶的活性各种外部信号(xìnhào)影响植物的生长发育光质→光受体→信号转导组分→光调节基因→向光性反应第二页,共70页。对于植物细胞来讲,有来自相邻细胞的刺激、细胞壁的刺激、激素、温度(wēndù)、光照等等刺激,连接环境刺激到植物反应的分子途径就是信号转导途径,细胞接受信号并整合、放大信号,最终引起细胞反应这种信息在胞间传递和胞内转导过程称为植物体内(tǐnèi)的信号传导第三页,共70页。植物的信号(xìnhào)分子信号传导分子途径:胞间信号传递(chuándì)膜上信号转换胞内信号转导(蛋白质可逆磷酸化)细胞反应。图6-25细胞信号传导的主要分子途径IP3.三磷酸肌醇;DG.二酰甘油(ɡānyóu);PKA.依赖cAMP的蛋白激酶;PKCa2+依赖Ca2+的蛋白激酶;PKC.依赖Ca2+与磷脂的蛋白激酶;PKCa2+·CaM.依赖Ca2+·CaM的蛋白激酶从而使细胞作出反应。胞间信号分子胞内信号分子按作用范围分:第四页,共70页。胞间信号(xìnhào)传递膜上信号(xìnhào)转换胞内信号转导胞内分子(fēnzǐ)反应第五页,共70页。一、胞间信号(xìnhào)细胞感受刺激后合成并传递到作用部位引起生理反应的化学物质。植物激素是植物体主要的胞间化学信号。如当植物根系受到水分亏缺胁迫时,根系细胞迅速合成脱落酸(ABA),ABA再通过木质部蒸腾流输送到地上部分,引起叶片生长(shēngzhǎng)受抑和气孔导度的下降。而且ABA的合成和输出量也随水分胁迫程度的加剧而显著增加。这种随着刺激强度的增加,细胞合成量及向作用位点输出量也随之增加的化学信号物质称之为正化学信号(positivechemicalsignal)。然而在水分胁迫时,根系合成和输出细胞分裂素(CTK)的量显著减少,这样的随着刺激强度的增加,细胞合成量及向作用位点输出量随之减少的化学信号物质称为负化学信号(negativechemicalsignal)。植物体内(tǐnèi)的胞间信号可分为两类,即化学信号和物理信号。(一)化学信号
(chemicalsignals)干旱ABACTK正化学信号负化学信号第六页,共70页。当植物的一张叶片被虫咬伤后,会诱导本叶和其它叶产生蛋白酶抑制物(PIs)等,以阻碍病原菌或害虫进一步侵害。如果伤害后立即除去受害(shòuhài)叶,则其它叶片不会产生PIs。虫咬寡聚糖但如果将受害叶的细胞壁水解片段(主要是寡聚糖)加到叶片中,又可模拟伤害反应诱导PIs的产生,从而认为寡聚糖是由受伤叶片释放并经维管束转移,继而诱导能使PIs基因活化的化学信号(xìnhào)物质。虫咬不会(bùhuì)产生PIs产生PIs产生PIs第七页,共70页。受伤(shòushāng)西红柿植株蛋白激酶特制物生物合成快速诱导信导途径的假定模式图第八页,共70页。已知1,3-β-D-葡聚糖、寡聚半乳糖醛酸、富含甘露糖的糖蛋白、聚氨基葡萄糖等都是构成细胞壁的主要成分,它们除了具有(jùyǒu)支持细胞框架的功能外,还起诱导抗性和控制发育的信号作用,成为引人注目的胞间信号分子。此外,一些生长调节物质如壳梭孢菌素、花生四烯酸以及乙酰胆碱等也都具有(jùyǒu)化学信号的功能。
植物激素是植物体主要的胞间化学(huàxué)信号9大类植物(zhíwù)激素第九页,共70页。(二)物理(wùlǐ)信号(physicalsignal)指细胞感受到刺激后产生的能够起传递信息作用的电信号和水力学信号。电信号传递是植物体内长距离传递信息的一种重要方式,是植物体对外部刺激的最初反应。植物的电波研究较多的为动作电波(actionpotential,AP),也叫动作电位,它是指细胞和组织中发生的相对于空间和时间的快速变化的一类生物电位。植物中动作电波的传递仅用短暂的冲击(如机械震击、电脉冲或局部温度的升降)就可以激发出来,而且受刺激的植物没有伤害,不久便恢复原状。一些敏感植物或组织(如含羞草的茎叶、攀缘植物的卷须等),当受到外界(wàijiè)刺激,发生运动反应(如小叶闭合下垂、卷须弯曲等见录像)时伴有电波的传递。第十页,共70页。受触及的含羞草小叶在1至2秒钟向下弯,这是由于电波引发(yǐnfā)叶枕运动细胞中大量的K+和Ca+2转运,引起膨压改变的结果第十一页,共70页。图17.14Albiziapulvini背侧和腹侧的运动细胞之间的离子流调节(tiáojié)了小叶的开放与闭合。
第十二页,共70页。怀尔登(Wildon)等用番茄做实验,指出由子叶伤害而引起第一真叶产生蛋白酶抑制物PIs的过程中,动作电位是传播的主要方式。他们采取让电信号通过后马上就除去子叶以及使子叶叶柄致冷以阻碍筛管运输、排除化学物质传递的试验,其结果都证明单有电信号就可以引起PIs反应,而且(érqiě)他们也首次证明了电信号可引起包括基因转录在内的生理生化变化。植物细胞对水力学信号(压力势的变化)很敏感。玉米叶片木质部压力的微小变化就能迅速影响叶片气孔的开度,即压力势降低(jiàngdī)时气孔开放,反之亦然。伤害(shānghài)产生PIs电信号通过后去子叶第十三页,共70页。(三)胞间信号(xìnhào)的传递1.化学信号的传递(1)气相中传递易挥发性化学信号可通过植株体内的气腔网络扩散而迅速传递,传递速度可达2mm·s-1左右。乙烯和茉莉酸甲酯均属此类信号。(2)韧皮部传递植物体内许多化学信号物质,如IAA、茉莉酸甲酯、寡聚半乳糖、水杨酸等都可通过韧皮部途径传递。(3)木质部传递化学信号可通过集流的方式在木质部内传递。土壤干旱胁迫时,根系可迅速合成并输出ABA。合成的ABA可通过木质部蒸腾流进入叶片,并影响叶片中的ABA浓度,从而抑制叶片的生长和气孔的开放(kāifàng)。3.电信号的传递植物电波信号的短距离传递需要通过共质体和质外体途径,而长距离传递则是通过维管束。4.水力学信号的传递水力学信号是通过植物体内水连续体系中的压力变化来传递的。第十四页,共70页。RobertF.FurchgottSUNYHealthScienceCenter
Brooklyn,NY,USA.1916~
LouisJ.IgnarroDept.ofMolecularandMedicalPharmacology
UCLASchoolofMedicine
LosAngeles1941~FeridMuradDept.ofIntegrativeBiology
PharmacologyandPhysiology
UniversityofTexasMedicalSchool,HoustonANewPrincipleNitricOxide,NO,isashort-lived,endogenouslyproducedgasthatactsasasignalingmoleculeinthebody.Signaltransmissionbyagas,producedbyonecell,whichpenetratesmembranesandregulatesthefunctionofothercellsisanentirelynewprincipleforsignalinginthehumanorganism.NOasasignallingmoleculefor1998Nobelprize
第十五页,共70页。Furchgott'ssandwichRobertFFurchgottshowedthatacetylcholine-inducedrelaxationofbloodvesselswasdependentontheendothelium.His"sandwich"experimentsetthestageforfuturescientificdevelopment.Heusedtwodifferentpiecesoftheaorta;onehadtheendotheliallayerintact,intheotherithadbeenremoved.第十六页,共70页。Hemoglobin(yellow)exposedtoendothelialcellsthatwerestimulatedtoproduceEDRF(green)
Hemoglobin(yellow)directlyexposedtoNO(green)Theshiftofabsorptioncurvesisidentical,henceEDRFisNOIgnarro'sspectralanalysisLouisIgnarroreportedthatEDRFrelaxedbloodvessels.HealsoidentifiedEDRFasamoleculebyusingspectralanalysisofhemoglobin.WhenhemoglobinwasexposedtoEDRF,maximumabsorbancemovedtoanewwave-length;andexposedtoNO,exactlythesameshiftinabsorbanceoccurred!EDRFwasidenticalwithNO.Anewprincipleforsignallingbetweenhumancellswasdiscovered.第十七页,共70页。Murad'senzymeactivationFeridMuradknewthatnitroglycerinecausedrelaxationofsmoothmusclecells.Theenzyme,guanylylcyclase,wasactivatedandincreasedcyclicGMP,causingrelaxationofthemuscle.Didnitroglycerinactviareleaseofnitricoxide,NO?HebubbledNO-gasthroughtissuecontainingtheenzyme;cyclicGMPincreased!Anewmodeofdrugactionhadbeendiscovered!第十八页,共70页。Inatherosclerosis,plaquesreducebloodflowinthearteries.Thisdecreasesoxygensupplytotheheartmusclecausingchestpain(anginapectoris)andsometimesevenmyocardialinfarction.TreatmentwithnitroglycerineprovidesNO,dilatesthevessels,andincreasesbloodflow.Thankstothisyear'sNobelLaureateswenowunderstandhownitroglycerine,animportantheartmedicine,works.ItactsasaNOdonor,causesdilationofthebloodvessels,increasesoxygensupplyandprotectstheheartfromdamageandcelldeath.Nitroglycerinea100yearoldexplosiveandheartmedicine第十九页,共70页。AlfredNobelsufferedfromanginapectorisandwasprescribednitroglycerine.Inalettertoafriendhewrote:"ItsoundsliketheironyoffatethatIhavebeenprescribednitroglycerineinternally.TheyhavenameditTrinitrininordernottoupsetpharmacistsandthepublic.
Youraffectionatefriend,
A.Nobel"NOisimportantforsignallingbetweennervecellsinthebrain
NOisinvolvedinthenormaldefenceagainstbacterialandparasiticinfections
第二十页,共70页。二、膜上信号(xìnhào)的转换胞间信号从产生位点经长距离传递(chuándì)到达靶细胞,靶细胞首先要能感受信号并将胞外信号转变为胞内信号,然后再启动下游的各种信号转导系统,并对原初信号进行放大以及激活次级信号,最终导致植物的生理生化反应。信号转导:刺激→受体→信号整合(zhěnɡhé)与放大→基因表达→反应第二十一页,共70页。(一)受体与信号(xìnhào)的感受受体(receptor)是指在效应器官细胞质膜上或亚细胞组分中能与信号物质特异性结合,并引发产生胞内次级信号的特殊成分。与能受体结合的特殊信号物质称配体(Ligang)。受体可以(kěyǐ)是蛋白质,也可以(kěyǐ)是一个酶系。受体的主要特性:①能与配体特殊(tèshū)结合;②高度的亲和力;③饱和性。根据受体在细胞中的位置,可将它分为细胞表面受体和胞内受体。第二十二页,共70页。外部信号与细胞表面的或与细胞质或核内部的受体结合。许多(xǔduō)亲水的分子如肽、碳水化合物和渗透的信号不易通过质膜,因而在细胞表面被察觉(如配体1),两性的和疏水的分子,如生长调节剂,能通过质膜,因而能被细胞表面受体或细胞内部的受体察觉(如配体2)在多数情况下信号分子与细胞表面的受体分子结合。许多亲水的分子如肽、碳水化合物和渗透的信号不易通过质膜,而与细胞表面的受体相结合一般认为受体存在于质膜上。然而植物细胞具有细胞壁,它可能使某些胞间信号分子不能直达膜外侧,而首先作用于细胞壁。一些(yīxiē)外界刺激有可能通过细胞壁—质膜—细胞骨架蛋白变构而引起生理反应。第二十三页,共70页。两性的和疏水的分子,如生长调节剂,能通过质膜,因而能被细胞表面受体或细胞内部(如细胞核上)的受体结合。甾类化合物是疏水性小分子,可以简单(jiǎndān)扩散进入细胞,与细胞内受体结合,调节基因转录。目前研究比较活跃(huóyuè)的受体是光敏受体和激素受体以及可能起受体作用的激发子结合蛋白。茉莉酸的生物合成(héchéng)和信号转导示意图13S-HPOT.13-过氧羟基亚麻酸;12-OXO-PDA.12-氧代-植物二烯酸;OPC-8∶0.3氧代-(2′-戊烯基)-环戊烷-1-辛酸第二十四页,共70页。18.4蓝光和红光经常相互影响和交替作用于植物的发育.DET(去黄化)和COP(光形态(xíngtài)发生组分)两类蛋白通常维持暗中黄化过程,第二十五页,共70页。(A)G蛋白偶连受体活化时G蛋白连接受体传递信息到G蛋白质,其上有GTP。GTP复合体中的α-亚基能与β、γ亚基分开,进入细胞质激活其他酶。(B)酶偶连受体受体通常是蛋白激酶,与信号结合(jiéhé)后,随受体活化,内部分子磷酸化,传递信息。(C)离子通道偶连受体受体可能本身细胞表面重要的通道。接受信号时,通道开放。也有些离子通道连接受体是在内部膜上。动物细胞中质膜上的三种(sānzhǒnɡ)类型的受体第二十六页,共70页。(二)G蛋白(dànbái)(Gprotein)在受体接受胞间信号分子到产生胞内信号分子之间,通常认为是通过G蛋白将信号转换偶联起来,故G蛋白又称偶联蛋白或信号转换蛋白。G蛋白全称为GTP结合调节蛋白(GTPbindingregulatoryprotein),此类蛋白由于其生理活性有赖于三磷酸鸟苷(GTP)的结合以及具有(jùyǒu)GTP水解酶的活性而得名。二十世纪70年代初在动物细胞中发现了G蛋白的存在,进而证明了G蛋白是细胞膜受体与其所调节的相应生理过程之间的主要信号转导者。G蛋白的信号偶联功能是靠GTP的结合或水解产生的变构作用完成。当G蛋白与受体结合而被激活时,继而触发效应器,把胞间信号转换成胞内信号。而当GTP水解为GDP后,G蛋白就回到原初构象,失去转换信号的功能。G蛋白的发现是生物学一大成就。吉尔曼(Gilman)与罗德贝尔(Rodbell)因此获得1994年诺贝尔医学生理奖。第二十七页,共70页。植物G蛋白的研究始于80年代,并已证明G蛋白在高等植物中的普遍存在,也证明它在光、植物激素对植物的生理效应、在跨膜离子运输、气孔运动(yùndòng)、植物形态建成等生理活动的细胞信号转导过程中有重要调节作用。小G蛋白与异源三体G蛋白α亚基有许多相似之处。它们都能结合(jiéhé)GTP或GDP,结合(jiéhé)了GTP之后都呈活化态,可以启动不同的信号转导。G蛋白(dànbái)细胞内的G蛋白一般分为两大类:异源三体G蛋白:由三种亚基(α、β、γ)构成
小G蛋白:只含有一个亚基的单体第二十八页,共70页。图18.30与GTP结合的人类Ras的鸟嘌呤核苷酸区域三维结构图;这种结构是用一种不可水解的GTP类似物测定的。被标记的10个环,将6个β折叠(箭头)与5个α螺旋连接起来。这种Ras结构是在动物,植物与真菌中发现的,其它(qítā)小分子GTP酶的模型。第二十九页,共70页。G蛋白位于膜内侧,并与质膜紧密结合。某种刺激信号与其膜上的特异受体结合后,激活的受体将信号传递给G蛋白,G蛋白的α亚基与GTP结合而被活化。活化的α亚基与β和γ亚基复合体分离而呈游离状态,活化的α亚基继而(jìér)触发效应器(如磷酸脂酶C)把胞外信号转换成胞内信号。而当α亚基所具有的GTP酶活性将与α亚基相结合的GTP水解为GDP后,α亚基恢复到去活化状态并与β和γ亚基相结合为复合体。这样完成一次循环。
18.31异源三体G蛋白的活动(huódòng)循环1234第三十页,共70页。图18.36磷脂酶A2(PLA2)在植物信号反应中假定的作用。PLA2的活化作用将磷脂酰胆碱(PC)水解成为溶血卵磷脂(LysoPC)释放的自由(zìyóu)脂肪酸可以用于调节基因表达诱导作用的茉莉酸。虽然连接所有组分的确切关系尚未完全阐明,这种途径也无疑在胁迫信号的传导过程中具有重要的作用。第三十一页,共70页。如果将胞外各种刺激信号作为(zuòwéi)细胞信号传导过程中的初级信号或第一信使,那么则可以把由胞外刺激信号激活或抑制的、具有生理调节活性的细胞内因子称细胞信号传导过程中的次级信号或第二信使(secondmessenger)。胞外信号经跨膜转换之后,通过第二信使进一步传递和放大,最终引起细胞内的生化反应。三、胞内信号(xìnhào)的转导第三十二页,共70页。植物(zhíwù)中的的第二信使,主要是cAMP、cGMP、DAG、IP3、Ca2+等。二酰甘油(ɡānyóu)肌醇-1,4,5-三磷酸(línsuān)第三十三页,共70页。植物细胞质中Ca2+含量一般在10-7~10-6mol·L-1,而胞外Ca2+浓度约为10-4~10-3mol·L-1;胞壁是细胞最大的Ca2+库,其浓度可达1~5mol·L-1;细胞器的Ca2+浓度也是胞质的几百到上千倍,所以Ca2+在植物细胞中的分布(fēnbù)极不平衡。1.钙信号系统钙离子植物细胞内的游离钙离子是细胞信号转导过程(guòchéng)中重要的第二信使第三十四页,共70页。植物细胞内Ca2+存储之间的关系是复杂的,Ca2+浓度在细胞器中是高的,但在细胞壁中与细胞质中是低的。当细胞发生信号反应时,多种细胞器上或质膜上的通道都打开了,使得Ca2+能够沿着其化学梯度扩散进入细胞质。Ca2+ATP酶以及可能Ca2+/H+反向运输载体将细胞质浓度返回到其开始的数值。所知的是,显示(xiǎnshì)的Ca2+的亚细胞浓度(内质网所引用的值从0.1到1mM不等)。细胞质结合位点的浓度已经在约0.5到1mM时测量。自由的细胞质Ca2+与这些结合位点是相等的。细胞质Ca2+浓度的上升,活化了钙调素,因此(yīncǐ)启动了接下来的下游反应。IP3,1,4,5-三磷酸肌醇。图18.43细胞信号反应过程(guòchéng)中胞内外Ca2+的相互作用第三十五页,共70页。
质膜与细胞器上的Ca2+泵和Ca2+通道,控制细胞内Ca2+的分布和浓度;质膜上Ca2+泵将膜内的钙泵出细胞,质膜上Ca2+通道控制Ca2+内流,细胞器膜的Ca2+泵将胞质中Ca2+的积累在细胞器(胞内钙库)中,Ca2+通道则控制Ca2+外流。胞内外信号可调节这些(zhèxiē)Ca2+的运输系统,引起Ca2+浓度变化。植物(zhíwù)细胞中Ca2+的运输系统胞内游离钙离子(lízǐ)浓度的变化可能主要是通过钙离子(lízǐ)的跨膜运转或钙的螯合物的调节而实现的,此外,在质膜、液泡膜、内质网膜上都有钙离子(lízǐ)泵或钙离子(lízǐ)通道的存在(图6-26)第三十六页,共70页。许多信号如蓝光、触摸能改变膜势和活化通道,使钙能进入,增加胞质中Ca+,这样(zhèyàng)引起K+和Cl+通道的打开,失去膨压,这种分别的转导途径通过Ca+/钙调蛋白激酶和其他蛋白联系起来。蓝光和触摸也能通过其他路线改变细胞内的Ca+、几乎所有的胞外刺激信号(如光照(guāngzhào)、温度、重力、触摸等物理刺激和各种植物激素、病原菌诱导因子等化学物质)都可能引起胞内游离钙离子浓度的变化,而这种变化的时间、幅度、频率、区域化分布等不尽相同,所以有可能不同刺激信号的特异性可靠钙离子浓度变化的不同形式来体现。第三十七页,共70页。细胞质中开放的Ca+2通道(tōngdào)附近Ca+2的分配颜色区表示Ca+2浓度,红的最高,蓝的最低第三十八页,共70页。红光诱导小麦叶片原生质体胞质中Ca2+的变化,用对Ca2+敏感的螢光染料-3,共焦显微图。高Ca2+处为红色,低Ca2+处为蓝色。数字为加入染料后时间(shíjiān)秒,只能看到一半的原生质体,因为螢光染料不能进入液泡。第三十九页,共70页。图18.48花粉管在其顶端区域维持了一个稳定的细胞质Ca2+梯度。[Ca2+]i的稳定梯度对生长尤为重要,是来源于与顶端相关的Ca2+通道(tōngdào)的聚焦。花粉管可以用例如indo-1或fura-2的Ca2+敏感性比例的成像染色剂,并通过荧光显微照相相对于Ca2+进行定量。第四十页,共70页。图18.52鸭跖草单个保卫细胞在0时刻将其在20μM到1mM的培养介质中改变其Ca2+浓度后,[Ca2+]的荧光比例图象。单个保卫细胞用比例染色剂indo-1进行染色。在介质改变之后的(从左向右)0,2与5分钟(上排)以及10,20,30分钟(下排)后所拍摄的比例照片(zhàopiàn)。明亮区域的形象是插入来表现保卫细胞的气孔。第四十一页,共70页。图18.53(A)细胞质中Ca2+的摆动可能是来源于内质网Ca2+储存的充实与空虚。箭头的大小表示了流动的速率(sùlǜ)。当内质网缺乏Ca2+时,Ca2+被螯合进入了内质网。当内质网Ca2+充实的时候,Ca2+被释放出来。这两种机理的基础可能是通过细胞器中的通道与ATP酶活性的改变起作用。(B)[Ca2+]i的摆动伴随着花粉管的生长而进行。荧光比例图象是以1分钟间隙拍摄,从而揭示了尖端[Ca2+]的摆动。第四十二页,共70页。图18.54生长花粉管中的Ca2+波(A)罂粟花粉管在Ca2+-敏感性的荧光染色剂,fluo-3的彩色图象,这些图象表现的是0时刻(图片的顶部)处理的1,4,5-三磷酸肌醇光解后的情况。诱导的Ca2+波是在细胞(xìbāo)核的附近(在花粉管细胞(xìbāo)质的中央区域)。这些波大约在1分钟内到达花粉管的顶端,在其前进的过程中,可能会摆动。这些照片是在光解释放后特定的时间间隔下拍摄的。(B)在束缚态IP3光解作用后,在花粉管中可以观察到Ca2+的演绎机制。此波极有可能通过质膜中Ca2+依赖的磷酸脂酶C而传播的,因为IP3的合成从细胞(xìbāo)内存储中动员了Ca2+。当波到达尖端时,生长便受到抑制;当其恢复时,生长就被重新定位了。第四十三页,共70页。如保卫细胞质膜上的内向(nèixiànɡ)钾离子通道可被钙离子抑制,而外向钾离子和氯离子通道则可被钙离子激活等。胞外刺激信号可能直接或间接地调节这些钙离子(lízǐ)的运输系统,引起胞内游离钙离子(lízǐ)浓度变化以至影响细胞的生理生化活动。第四十四页,共70页。图3.49Ca2+信号反应在气孔关闭过程中,与多个离子(lízǐ)通道的活性及H+泵相协调①受体(R)感受ABA的作用导致了Ca2+的输入或Ca2+从内部储存中的释放,②细胞质中的自由Ca2+浓度提高↑,促进了质膜上阴离子(lízǐ)与K+Out通道的开放,并抑制了K+in通道的开放。③离开细胞的离子(lízǐ)比进入细胞的多时,流出细胞水的量就会超过进入的水,如此,细胞就会失去膨胀作用,从而使得气孔关闭。第四十五页,共70页。图18.26保卫细胞中束缚态ABA的光解会导致气孔关闭。(A)一种束缚态的ABA,1-(2-硝基苯基)-乙基-ABA,它是依赖于UV光的。当其被装载进入保卫细胞并被光分解(fēnjiě)时,束缚态的ABA就会转变为自由态的ABA,并导致保卫细胞的关闭。(B)表现ABA对气孔效应的光学显微照片。保卫细胞被用束缚态的ABA进行装载(左图)并发生光裂解,从而导致了气孔的关闭(右图)。第四十六页,共70页。图18.27ABA可能的传导途径。通常认为ABA与一个(yīɡè)受体相结合(未显示),从而允许了质膜上Ca2+的直接进入并激发了一种Ca2+敏感的蛋白质磷酸酶(ABI1)。第四十七页,共70页。胞内Ca2+信号通过(tōngguò)其受体-钙调蛋白转导信号。现在研究得较清楚的植物中的钙调蛋白主要有两种:钙调素和钙依赖型蛋白激酶。钙调素(calmodulin,CaM)是最重要的多功能Ca2+信号受体,由148个氨基酸组成的单链的小分子(分子量为17000~19000)酸性蛋白。CaM分子有四个Ca2+结合位点。当外界信号刺激引起胞内Ca2+浓度上升到一定阈值后(一般≥10-6mol),Ca2+与CaM结合,引起CaM构象改变。而活化的CaM又与靶酶结合,使靶酶活化而引起生理反应。目前已知有十多种酶受Ca2+-CaM的调控(diàokònɡ),如多种蛋白激酶、NAD激酶、H+-ATPase、Ca2+-ATP酶、Ca2+通道等。在以光敏色素为受体的光信号传导过程中Ca2+-CaM的信号系统也起着重要的调节作用。生长素、光、摩擦等都可引起CaM基因活化,使CaM含量增加。钙调素第四十八页,共70页。A.钙调素呈哑铃形,长为6.5nm,每个哑铃球是有2个Ca2+结合位点,长的中心螺旋形成(xíngchéng)哑铃柄,无Ca2+结合时,两个球部沿着中心螺旋折叠。B.形成(xíngchéng)Ca2+-CaM复合体后,结合到靶酶上。Ca2+-CaM复合体的形成(xíngchéng)使CaM与许多靶酶的亲和力大大提高。第四十九页,共70页。蔡南海实验室用光敏色素A缺失的番茄单细胞突变系,研究了完整叶绿体中多种光系统色素蛋白质及酶基因(jīyīn)到达所必需的光信号传导过程。将信号系统中各组分,如phyA、G蛋白激活剂,Ca2+及CaM、cGMP、cAMP等以及它们的不同组合微注射入单细胞内,随后对叶绿体内多种色素蛋白和酶系在基因(jīyīn)活化、转录、翻译三个水平进行观察缺少光敏色素番茄突变体用微量注射纯的光敏色素后用红光照射可促使叶绿体发育。这一处理效果(xiàoguǒ)可被微量注射cGMP和Ca2+替代。第五十页,共70页。研究后发现:光刺激后,上游的光敏色素与G蛋白(dànbái)激活对完整叶绿体发育及花色素苷合成是共同必须的;Ca2+-CaM系统参与PSⅡ捕光色素、ATP合成酶、Rubisco等光诱导产生,但不参与诱导花色素苷合成,因而只形成不成熟的叶绿体;而在动物视觉系统细胞光感应中起重要作用的cGMP在植物花色素苷诱导中起决定作用,并与Ca2+-CaM一起诱导PSⅠ和Cytb6/f的合成(图6-27)。图6-27光诱导的花色素苷和叶绿体某组分(zǔfèn)的合成第五十一页,共70页。2.肌醇磷脂(línzhī)信号系统对于生物膜在信号转导中的作用,多年来人们把注意力集中在功能(gōngnéng)繁多的膜蛋白上,而脂质组分仅被看成是种惰性基质。80年代后期的研究表明,质膜中的某些磷脂,在植物细胞内的信号转导过程中起了重要作用。肌醇磷脂是一类由磷脂酸与肌醇结合的脂质化合物,分子中含有甘油、脂酸磷酸和肌醇等基团,其肌醇分子六碳环上的羟基被不同数目的磷酸酯化,其总量约占膜磷脂总量的1/10左右,主要以三种形式存在于植物质膜中,即:磷脂酰肌醇(PI)、磷脂酰肌醇-4-磷酸(PIP)磷脂酰肌醇-4,5-二磷酸(PIP2)。PIPIPPIP2第五十二页,共70页。特定(tèdìng)的磷脂酶在特定(tèdìng)的分裂位点降解膜磷脂图6-28肌醇磷脂的分子结构箭头所示为不同(bùtónɡ)磷脂酶(A1、A2、C、D)的作用位点,肌醇环上4,5位羟基可在专一性激酶作用下磷酸化第五十三页,共70页。细胞外刺激(cìjī)信号或激素等物质被质膜上专一受体接受,经G-蛋白激活磷酸脂酶C(PLC),使磷脂酰肌醇-4,5-二磷酸(PIP2)水解生成二酰甘油(DAG)和肌醇-1,4,5-三磷酸(IP3),IP3与细胞内贮钙体膜上专一受体结合,促使Ca2+从液泡中释放出来,Ca2+与CaM结合形成Ca2+-CaM复合物,仍留在质膜上的DAG使蛋白激酶C(PKC)激活,催化蛋白质磷酸化;Ca2+-CaM和蛋白质(酶)磷酸化导致细胞反应。IP3产生后很快被磷酸二酯酶水解,首先产生肌醇-1,4-二磷酸(IP2),继而是肌醇单磷酸(IP)及磷脂酰肌醇(PI),返回膜中。⊕表示活化。肌醇磷酸代谢(dàixiè)循环过程IP3作为(zuòwéi)信号分子,在植物中它的主要作用靶为液泡,IP3作用于液泡膜上的受体后,可影响液泡膜形成离子通道,使Ca2+从液泡这一植物细胞中重要的贮钙体中释放出来,引起胞内Ca2+浓度提高,从而启动胞内Ca2+信号系统来调节和控制一系列的生理反应。DAG的受体是蛋白激酶C(PKC),在一般情况下,质膜上不存在自由的DAG。在有DAG、Ca2+时,磷脂与PKC分子相结合,PKC激活,使某些酶类磷酸化,导致细胞反应;第五十四页,共70页。由磷酸脂酶C的活化到引起胞质钙增加的传导顺序。与质膜结合的磷酸脂酶C由G蛋白活化,磷脂酰-4,5-二磷酸由磷酸脂酶C水解产生第二信使肌醇-1,4,5-三磷酸和二酰甘油,肌醇-1,4,5-三磷酸活化与内质网或液泡连结(liánjié)的肌醇-1,4,5-三磷酸受体,而开始释放Ca2+。磷脂化合物磷脂酰肌醇、磷脂酰肌醇-4-磷酸、肌醇-1,4-二磷酸和肌醇单磷酸循环使用。肌醇-1,4-二磷酸肌醇单磷酸磷脂(línzhī)酰肌醇肌醇-1,4,5-三磷酸(línsuān)磷脂酰-4,5-二磷酸二酰甘油磷脂酰肌醇-4-磷酸磷酸脂酶C第五十五页,共70页。图18.37在磷脂酶C(PLC)与磷脂酶D(PLD)之间可能相互作用的推测图。磷脂酸(PtdOH)从膜磷脂因为PLD的作用而释放会诱导包含Ca2+与其它靶物(如,蛋白质激酶,肌动蛋白的组装与分泌)在内的几种信号反应的变化。多种因素,包括通道开放,异型三聚体G蛋白的促效及刺激的活化作用,以及由于热,冷或寄生虫而造成的膜的破坏,都能够造成膜的破坏,都能够通过钙调素结合区域(CalB)与自由Ca2+而活化PLD。Mastoparan(蜂毒中的一种多肽),霍乱病毒以及乙醇(yǐchún)能够直接模仿G-蛋白的活化作用。活化的PLD由重新回到质膜上,并在此水解了磷脂酰胆碱(PC)。PtdOH可以作为离子载体,使得更多的Ca2+进入细胞质。
PtdOH同时也可以一个由DAG激酶催化的反应中的甘油二脂(DAG)合成。伴随的DAG相关的PLC的激发作用将会进一步提高DAG激酶的活性。PtdOH也可能会由于PtdOH激酶作用的甘油二脂焦磷酸(DGPP)的转化作用而发生(fāshēng)钝化。PtdOH的任何增加都会扩大PLC的信号的级联反应,这个反应是通过激发磷脂酰激酶肌醇-4-磷酸-5-激酶(PIPKIN)反应而进行,这个激发反应将PIP转化为PLC底物PIP2。第五十六页,共70页。还有一些物质被认为具有第二信使的作用,如cGMP(环鸟苷酸)、H+、某些(mǒuxiē)氧化还原剂(如抗坏血酸、谷光甘肽和过氧化氢)。受动物细胞信号的启发,人们最先在植物中寻找的胞内信使是环腺苷酸(cyclicAMP,cAMP),但这方面的进展较缓慢,在动物细胞中,cAMP-依赖性蛋白激酶(蛋白激酶A,PKA)是cAMP信号系统的作用中心。环3’,5’-AMP(cAMP)的合成和分解分别由腺苷环化酶和环AMP磷酸二酯酶催化。植物中也可能存在着PKA。蔡南海实验室证实了在叶绿体光诱导花色素苷合成过程中,cAMP参与受体G蛋白之后的下游信号转导过程,环核苷酸信号系统与Ca2+-CaM信号转导系统在合成完整(wánzhěng)叶绿体过程中协同起作用(见下页图)。3.环核苷酸信号系统cAMP第五十七页,共70页。蔡南海(NamHaiChan)实验室用光敏色素A缺失(quēshī)的番茄单细胞突变系,研究了完整叶绿体中多种光系统色素蛋白质及酶基因表达所必需的光信号传导过程。将信号系统中各组分,如光敏色素A、G蛋白激活剂,Ca2+及CaM、cGMP、cAMP等以及它们的不同组合微注射入单细胞内,随后对叶绿体内多种色素蛋白和酶系在基因活化、转录、翻译三个水平进行观察研究后得出结论:光刺激后,上游的光敏色素与G蛋白激活对完整叶绿体发育及花色素苷合成是共同必须的;Ca2+-CaM系统参与PSⅡ捕光色素、ATP合成酶、Rubisco等光诱导产生,但不参与诱导花色素苷合成,因而只形成不成熟的叶绿体;而在动物视觉系统细胞光感应中起重要作用的cGMP在植物花色素苷诱导中起决定作用,并与Ca2+-CaM一起诱导PSⅠ和Cytb6/f的合成图6-27光诱导的花色素苷和叶绿体某组分(zǔfèn)的合成第五十八页,共70页。四、蛋白质的磷酸化和去磷酸化植物体内许多功能(gōngnéng)蛋白转录后需经共价修饰才能发挥其生理功能(gōngnéng),蛋白质磷酸化就是进行共价修饰的过程。蛋白质磷酸化以及去磷酸化是分别由一组蛋白激酶(proteinkinase)和蛋白磷酸酯酶(proteinphosphatase)所催化的,它们是上述的几类胞内信使进一步作用的靶酶,也即胞内信号通过调节胞内蛋白质的磷酸化或去磷酸化过程而进一步转导信号。蛋白质可逆磷酸化反应式NTP代表(dàibiǎo)ATP或GTP,NDP代表(dàibiǎo)ADP或GDP,第五十九页,共70页。外来信号与相应的受体结合,会导致后者构象发生变化,随后就可通过引起第二信使的释放(shìfàng)而作用于蛋白激酶(或磷酸酯酶),或者因有些受体本身就具有蛋白激酶的活性,所以与信号结合后可立即得到激活,引起细胞反应。
胞内信号通过调节胞内蛋白质的磷酸化或去磷酸化过程(guòchéng)而进一步转导信号第六十页,共70页。18.63许多由蛋白磷酸化酶传导(chuándǎo)的信号,级联反应和基因表达。MAPK-活化的促进细胞分裂的蛋白激酶TF-转录(zhuǎnlù)因子Ras-一种小的GTP酶通过受体激活一系列的蛋白激酶,导致蛋白质磷酸化的级联(cascade)反应,在一系列反应中,前一反应的产物是后一反应中的催化剂,每次修饰就产生一次放大(fàngdà)作用第六十一页,共70页。蛋白激酶可对其底物蛋白质所特定的氨基酸残基进行磷酸化修饰,从而引起相应的生理反应,以完成信号转导过程。此外,由于蛋白激酶的底物既可以是酶,也可以是转录因子,因而它们既可以直接通过对酶的磷酸化修饰来改变酶的活性,也可以通过修饰转录因子而激活或抑制基因的表达,从而使细胞对外来信号作出相应的反应。蛋白激酶是个大家族,植物中约有2%-3%的基因编码蛋白激酶。根据磷酸化靶蛋白的氨基酸残基的种类不同,蛋白激酶有丝氨酸/苏氨酸激酶、酪氨酸激酶和组氨酸激酶等3类,它们分别将底物蛋白质丝氨酸/苏氨酸、酪氨酸和组氨酸残基磷酸化。有的蛋白激酶具有双重底物特异性,既可使丝氨酸或苏氨酸残基磷酸化,又可使酪氨酸磷酸化。在植物中,目前已知的蛋白激酶至少有30多种,它们的作用(zuòyòng)也表现在多个方面,包括向光性、抗寒、抗病、根部的向地性、光合作用(zuòyòng)、自交不亲和性以及细胞分裂等。
(一)蛋白激酶第六十二页,共70页。蛋白激酶中研究得较多的是最初从大豆中得到的依赖于钙离子的蛋白激酶(calciumdependentproteinkinase,CDPK),CDPK有一与钙结合位点。一般来说,CDPK在其氨基端有一个(yīɡè)激酶活性域,在其羧基端有一个(yīɡè)类似CaM的结构区域,在两者之间还有一个(yīɡè)抑制域。当位于CDPK上类似CaM的结构区域上的钙离子结合位点与Ca2+结合后,抑制被解除,酶就被活化。大豆、玉米、胡萝卜、拟南芥等植物中都存在这类蛋白激酶。现已知的可被CDPK磷酸化的作用靶(或底物分子(fēnzǐ))有细胞骨架成分、膜运输成分、离子通道、质膜上的质子ATP酶等。钙依赖型蛋白激酶+Ca+2激酶(jīméi)活性第六十三页,共70页。如从燕麦中分离出与质膜成分相结合的CDPK成分可将质膜上的质子
温馨提示
- 1. 本站所有资源如无特殊说明,都需要本地电脑安装OFFICE2007和PDF阅读器。图纸软件为CAD,CAXA,PROE,UG,SolidWorks等.压缩文件请下载最新的WinRAR软件解压。
- 2. 本站的文档不包含任何第三方提供的附件图纸等,如果需要附件,请联系上传者。文件的所有权益归上传用户所有。
- 3. 本站RAR压缩包中若带图纸,网页内容里面会有图纸预览,若没有图纸预览就没有图纸。
- 4. 未经权益所有人同意不得将文件中的内容挪作商业或盈利用途。
- 5. 人人文库网仅提供信息存储空间,仅对用户上传内容的表现方式做保护处理,对用户上传分享的文档内容本身不做任何修改或编辑,并不能对任何下载内容负责。
- 6. 下载文件中如有侵权或不适当内容,请与我们联系,我们立即纠正。
- 7. 本站不保证下载资源的准确性、安全性和完整性, 同时也不承担用户因使用这些下载资源对自己和他人造成任何形式的伤害或损失。
最新文档
- 2024年篮球裁判员应急方案试题及答案
- 2024年模具设计师考试大纲的全面解读试题及答案
- 学习重点 2024年体育经纪人考试知识点分解试题及答案
- 智能运输物流中心项目可行性研究报告(范文参考)
- 深度解析2024年篮球裁判员试题及答案汇编
- CAD软件在模具设计中的萎缩试题及答案
- 2024年农业植保员考试复习深入试题及答案
- 卵巢储备功能下降中西医结合诊疗指南
- 组合学习模式助力注册会计师考试成功试题及答案
- 2024年模具设计师资格考试考点大解析试题与答案
- 3DMAX创建之美知到智慧树章节测试课后答案2024年秋郑州信息工程职业学院
- 2024届九省联考英语试题(含答案解析、MP3及录音稿)
- 医院新技术项目鉴定审批表
- 【MOOC】College Students'Innovation and Entrepreneurship Practice-Southwest Jiaotong University 中国大学慕课MOOC答案
- 2024年司法考试刑法真题及答案
- 2024年天津市高考化学试卷(含答案逐题解析)
- 《工程伦理》练习题集
- 大型活动策划与管理第十一章 大型活动后勤保障
- 港航实务 皮丹丹 教材精讲班课件 52-第2章-2.5.3-铺面面层施工-2.5.4-铺面连接施工-2.5.5-堆场构筑物施工
- 危险品仓储危险品贮运车辆考核试卷
- 酒店工作安全培训(共60张课件)
评论
0/150
提交评论