



版权说明:本文档由用户提供并上传,收益归属内容提供方,若内容存在侵权,请进行举报或认领
文档简介
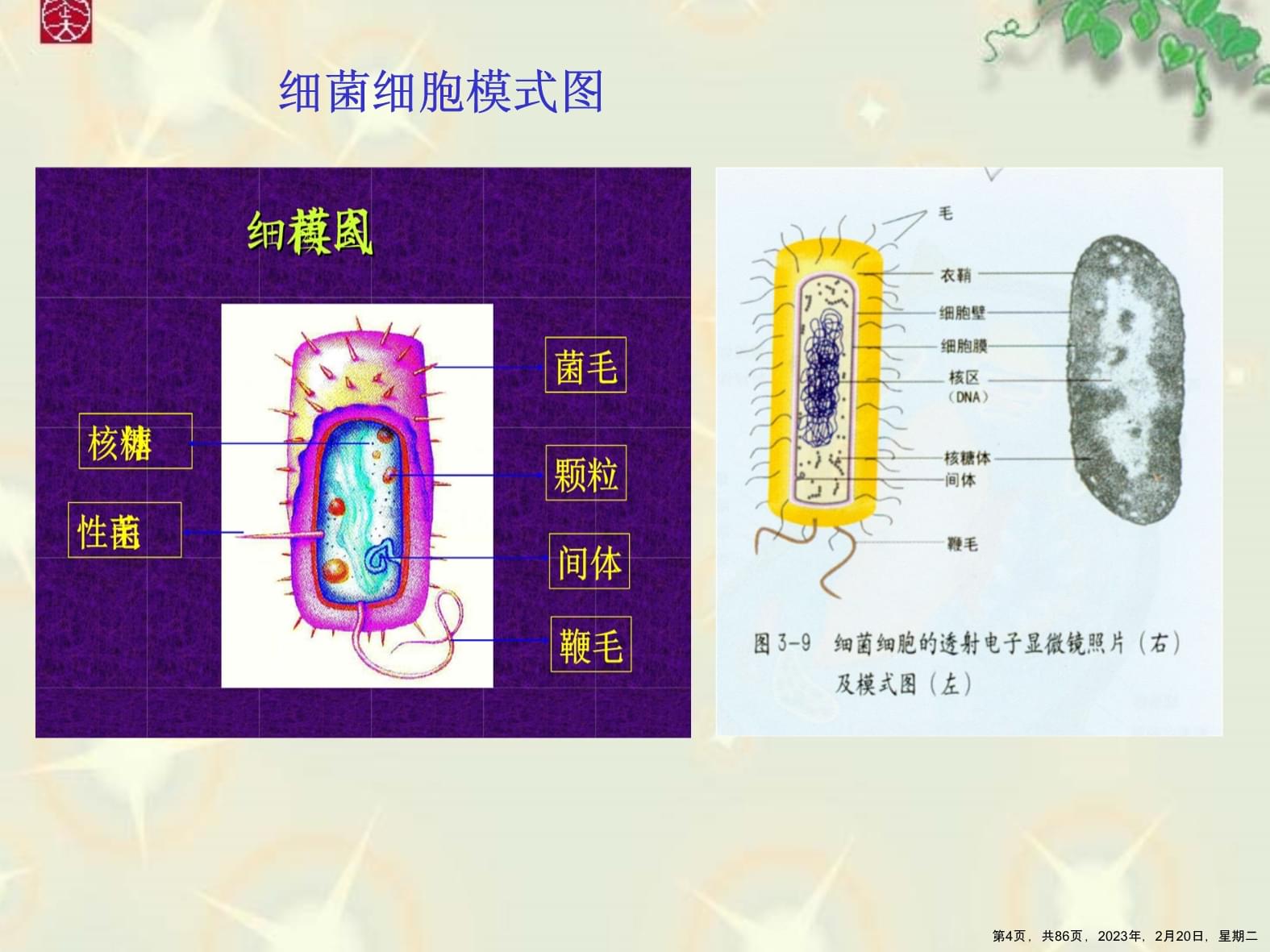
第十章细菌和病毒的遗传第1页,共86页,2023年,2月20日,星期二第一节细菌和病毒遗传研究的意义*一、细菌的生物学特征*二、病毒的生物学特征三、细菌和病毒在遗传研究中的优越性四、细菌和病毒的拟有性过程五、细菌遗传的实验研究方法第2页,共86页,2023年,2月20日,星期二一、细菌的生物学特征所有的细菌都是比较小的细胞,、大约1-2μm长,0.5μm宽,。由一层或多层膜或壁所包围。细菌细胞中有含有许多核糖体的细胞质,以及含DNA的区域,即拟核。大肠杆菌(Escherichiacoli)在细菌遗传学研究中应用十分广泛,其DNA主要是以单个主染色体(mainchromosome)的形式存在。第3页,共86页,2023年,2月20日,星期二细菌细胞模式图第4页,共86页,2023年,2月20日,星期二二、病毒的生物学特征病毒没有细胞结构,既不属于原核生物,也不属于真核生物。可分菌类病毒,动物病毒和植物病毒。病毒结构十分简单,仅含一种核酸,或DNA或RNA,和一个蛋白质外壳。蛋白质外壳保护遗传物质,并参与感染宿主细胞的过程。在病毒中没有合成蛋白质外壳所必须的核糖体。第5页,共86页,2023年,2月20日,星期二病毒的生物学特征病毒必须感染活细胞,改变和利用活细胞的代谢合成机器,才能合成新的病毒后代。感染细菌的病毒又叫噬菌体(bacteriophage)。依其遗传物质的性质,可将病毒分为单链DNA、单链RNA、双链DNA和双链RNA等四种类型。依其与宿主细胞的相互关系,可以有烈性(virulent)和温和(temperate)噬菌体两大类型。第6页,共86页,2023年,2月20日,星期二病毒的特性第7页,共86页,2023年,2月20日,星期二三、细菌和病毒在遗传研究中的优越性了解微生物遗传研究有助于理解多年来分子生物学、分子遗传学理论发展。世代周期短,繁殖世代所需时间短;易于操作管理和进行化学分析(纯培养与代谢产物累积);便于研究基因的突变(表现与选择);便于研究基因的作用(突变型生长条件与基因作用);便于研究基因的重组(重组群体大、选择方法简便有效);遗传物质比较简单,可作为研究高等生物的简单模型在研究基因结构、功能与表达调控时更为简便。微生物的应用领域日益扩大、成就突出(微生物工程);在遗传工程中,作为重要研究材料、工具,也具有决定性作用。第8页,共86页,2023年,2月20日,星期二四、细菌和病毒的拟有性过程真核生物基因分离、自由组合及连锁交换均通过有性过程实现。细菌和病毒均不存在严格意义上的有性过程。细菌细胞内除了染色体外还有一些寄生性复制因子(如噬菌体和质粒),它们可以在细胞间传递,并且形成细菌染色体间以及细菌染色体与核外遗传因子间的重组体。这种重组体结构类似于真核生物减数分裂过程中形成的重组体结构。拟有性过程:引起细菌、病毒间遗传物质转移与重组的过程。拟有性过程的存在是细菌、病毒在遗传学研究,特别是作为真核生物的模型研究遗传重组和基因结构的重要前提。第9页,共86页,2023年,2月20日,星期二五、细菌遗传的实验研究方法(一)细胞计数(培养物细胞浓度)(二)建立纯系的方法(三)选择培养法鉴定突变型与重组型(四)突变型与重组型的批量筛选方法第10页,共86页,2023年,2月20日,星期二(一)细胞计数(培养物细胞浓度)培养物中微生物计数方法是微生物学的基本实验技术,其基本思路是:对原培养物进行连续稀释;进行平板涂抹培养;由于每个细胞形成一个菌落,计数菌落数;根据稀释倍数计算原培养物中的细胞浓度。第11页,共86页,2023年,2月20日,星期二(二)建立纯系的方法——纯培养挑取由单个细胞繁殖而来的菌落进行培养就可以获得由一个细胞繁殖而来的纯系。通常采用平板表面涂布法或划线法可以获得单菌落。这种方法获得的纯系,称为“菌种纯”。有时采用显微操纵器进行菌丝尖端切割等方法从单个细胞直接培养建立纯系。采用这种方法获得的纯系称为“菌株纯”。第12页,共86页,2023年,2月20日,星期二(三)选择培养法鉴定突变型与重组型许多细菌的突变都与培养基营养成分及培养条件有关。营养缺陷型的筛选、鉴定:选择培养法是根据菌株在基本培养基和营养培养基上的生长表现将菌株分为原养型和营养缺陷型。营养突变型的筛选、鉴定方法与红色面包霉生化突变型的鉴定方法基本一致。其它突变类型的筛选、鉴定:对于其它的突变类型(如温度敏感型),也可以通过培养条件的选择培养来筛选与鉴定。第13页,共86页,2023年,2月20日,星期二(四)突变型与重组型的批量筛选方法为高效检测、分离混和群体中不同突变型,黎德伯格夫妇设计了影印培养法。该方法原理与选择培养法一致,但是采用影印法将在完全培养基上单菌落同时接种到不同选择培养基上同时对所有菌落进行选择培养,鉴定效率大大提高。注意:(1)最初的培养基必须是非选择性的,即各种突变型都能够在其上生长;(2)必须采用适当的方法如涂布或划线法,以使培养物菌落之间要分开。第14页,共86页,2023年,2月20日,星期二影印培养法第15页,共86页,2023年,2月20日,星期二细菌基因重组的方式一个细菌细胞的DNA与另一个细菌细胞DNA的交换重组可以通过四种不同的方式进行:转化接合性导转导第16页,共86页,2023年,2月20日,星期二第二节细菌的遗传分析一、转化二、接合三、性导第17页,共86页,2023年,2月20日,星期二一、转化(transformation)(一)、细菌转化实验(二)、转化过程(三)、共同转化与遗传图谱绘制第18页,共86页,2023年,2月20日,星期二转化(transformation)转化(transformation)是指某些细菌(或其他生物)能通过其细胞膜摄取周围供体(donor)的染色体片段,并将此外源DNA片段通过重组整合到自己染色体组的过程。只有当整合的DNA片段产生新的表现型时,才能测知转化的发生。转化中接受供体遗传物质的称为受体(receptor)。大部分的转化工作是用肺炎双球菌(Streptococcuspneumoniae)、枯草杆菌(Bacillussubtilis)和流感嗜血杆菌(Hemophilusinfluenzae)进行的。第19页,共86页,2023年,2月20日,星期二(一)、细菌转化实验1.基础知识野生型肺炎双球菌的菌落为光滑型,一种突变型为粗糙型,两者根本差异在于荚膜形成;荚膜的主要成分是多糖,具特殊的抗原性;不同抗原型是遗传的、稳定的,一般情况下不发生互变。荚膜菌落毒性类型光滑型S发达光滑有I,II,III粗糙型R无粗糙无I,II第20页,共86页,2023年,2月20日,星期二2.Griffith转化研究第21页,共86页,2023年,2月20日,星期二Griffith对其试验结论及发展根据上述研究结果Griffith认为:(有毒)死细菌中的某种物质转移到(无毒)活细菌中,并使之具有毒性,导致小家鼠死亡。他将这种细菌遗传类型的转变称为转化,并将引起转化的物质称为转化因子(tranformingprinciple)。以后一些研究者重复上述试验,并且加入了体外培养试验,即:将加热杀死的SIII细菌与无毒RII细菌混合培养,然后注入小家鼠体内,同样导致家鼠死亡。表明:细菌在培养条件下也能够实现遗传类型间的定向转化。第22页,共86页,2023年,2月20日,星期二3.阿维利(Avery)等的转化实验(1944)第23页,共86页,2023年,2月20日,星期二实验结论实验结果表明:来源于加热杀死的SIII细菌,并使RII细菌转化成为SIII型细菌的转化因子是DNA。转化定义为:某一基因型的细胞从周围介质中吸收来自另一基因型的DNA而使它的基因型和表现型发生相应变化的现象。Avery等人的实验也表明:决定细菌遗传类型的物质是DNA,即证明了DNA就是遗传物质。但由于他们采用的实验方法当时不被人们广泛接受,而且得出的结论与当时人们的传统观念(认为蛋白质是遗传物质)不符合,而长期没有得到人们的承认。第24页,共86页,2023年,2月20日,星期二(二)、转化过程不同细菌转化过程有一定差异,但是它们都存在几个共同特征:1.感受态与感受态因子:感受态指细菌能够从周围环境中吸收DNA分子进行转化的生理状态。感受态主要受一类蛋白质(感受态因子)影响,感受态因子可以在细菌间进行转移,从感受态细菌中传递到非感受态细菌中,可以使后者变为感受态。一般认为感受态出现在细菌对数生长后期,并且某些处理过程可以诱导或加强感受态,以大肠杆菌为例,用Ca2+(如CaCl2)处理对数生长后期的大肠杆菌可以增强其感受能力。第25页,共86页,2023年,2月20日,星期二转化过程2.供体(donor)DNA与受体(receptor)细胞结合(binding):结合发生在受体细胞特定部位;供体DNA片段为双链;结合过程是一个可逆过程。3.DNA的穿入和摄取:当细菌结合点饱和之后,细菌开始摄取外源DNA;往往只有一条DNA单链进入细胞,另一条链在膜上降解。4.联会(synapsis)与外源DNA片段整合(integration):整合就是指单链的转化DNA与受体DNA对应位点的置换,稳定地掺入到受体DNA中的过程。整合是一个遗传重组的过程。因而研究整合的分子机制也为遗传重组的分子机制作出了贡献。第26页,共86页,2023年,2月20日,星期二转化过程第27页,共86页,2023年,2月20日,星期二细菌转化过程第28页,共86页,2023年,2月20日,星期二(三)、共转化与遗传图谱绘制利用共同转化绘制细菌连锁遗传图谱的基本原理:相邻基因发生共同转化的概率与两者的距离间成反向关系,基因间距离越近,发生共同转化的频率越高,反之越低。因此可能通过测定两基因共同转化的频率来指示基因间的相对距离。第29页,共86页,2023年,2月20日,星期二转化与遗传图谱按照概率定律,两个片段同时转化的概率应为它们单独转化的概率的乘积,所以这种情况的概率是很低的。当两个基因紧密连锁时,它们就有较多的机会包括在同一个DNA片段中,并同时整合到受体染色体中。因此,紧密连锁的基因可以通过转化进行作图。例如,黎德伯格等人用枯草杆菌作了如下实验,即以trp2+his2+tyr1+为供体,以trp2-his2-tyr1-为受体进行转化,结果如表。
第30页,共86页,2023年,2月20日,星期二转化结果第31页,共86页,2023年,2月20日,星期二转化与遗传图谱可以看出,trp2、his2和tyr1是连锁的,其中his2和tyr1连锁紧密,这是因为它们并发转化的频率最高。根据重组值计算结果,可知trp2-his2的重组值为34%,trp2-tyr1的重组值为40%,his2-tyr1的重组值为13%,因此,trp2、his2及tyr1的排列顺序为:第32页,共86页,2023年,2月20日,星期二二、接合(conjugation)在原核生物中,接合是指遗传物质从供体—“雄性”转移到受体—“雌性”的过程。(一)、接合现象的发现和证实(二)、F因子及其在杂交中的行为(三)、中断杂交试验作图第33页,共86页,2023年,2月20日,星期二(一)、接合现象的发现和证实黎德伯格和塔特姆大肠杆菌杂交试验:材料:大肠杆菌(Escherichiacoli)K12菌株的两个营养缺陷型品系:A—甲硫氨酸缺陷型met-和生物素缺陷型bio-;B—苏氨酸缺陷型thr-和亮氨酸缺陷型leu-。方法:将A、B混和,在基本培养基(固体)上涂布培养。结果:平板上长出原养型菌落(++++)。第34页,共86页,2023年,2月20日,星期二黎德伯格和塔特姆接合试验第35页,共86页,2023年,2月20日,星期二几种可能解释及其分析对上述试验结果原养型菌落可能产生于:亲本细菌A或B发生了回复突变;两品系细胞通过培养基交换养料——互养作用;两品系间发生了转化作用;发生细胞融合,形成了异核体或杂合二倍体。为了验证这些原养型菌落产生的可能而进行的研究最终表明:这些解释均不成立。第36页,共86页,2023年,2月20日,星期二回复突变可能的排除Lederbery和Tatum利用的双营养缺陷型菌株进行试验,已基本排除A或B品系发生回复突变产生原养型细菌的可能。单基因回复突变的频率约为10-6;双基因回复突变的频率则为10-12,频率很低。但试验中产生原养型菌落产生的频率非常高,因此基本可以排除回复突变的可能。第37页,共86页,2023年,2月20日,星期二互养作用及其排除试验材料:A品系:A-B+T1S(met-bio-thr+leu+T1S);B品系:A+B-T1R(met+bio+thr-leu-T1R)。试验方法:将A、B品系混合接种在基本培养基表面;短时间后喷T1杀死A品系,使其不能持续产生thr与leu供B品系持续生长。结果与结论:仍然出现原养型菌落。从而表明互养并非原养型菌落出现的原因,而可能发生了遗传重组。第38页,共86页,2023年,2月20日,星期二转化作用及其排除把品系A的培养液经加热灭菌,加入到B品系的培养物中,未得到原养型菌落;表明原养型菌落可能不是由转化作用产生。戴维斯(Dawis,1950)的U型管试验(结果没有得到原养型细菌);实验结论:细胞直接接触是原养型细菌产生的必要条件。第39页,共86页,2023年,2月20日,星期二接合现象的发现和证实经过上述分析可以认为:在Lederbery和Tatum的试验中,发生了一种不同于转化的遗传重组方式,称之为接合。Hayes(1952)研究表明:大肠杆菌两种不同菌株(品系)接合过程中遗传物质的转移是单向的;从而认为大肠杆菌存在两种类型品系:雌性与雄性分别作为接合过程中遗传物质的供体与受体。接合(conjugation):遗传物质从供体(donor)转移到受体(receptor)的重组过程。第40页,共86页,2023年,2月20日,星期二接合现象的发现和证实第41页,共86页,2023年,2月20日,星期二(二)、F因子及其在杂交中的行为1.F因子:Hayes等进一步研究发现,大肠杆菌在接合中作供体的能力受细胞内一种致育因子(fertilityfactor,F因子;sexfactor,性因子)控制。F因子的化学本质是DNA,可以自主状态存在于细胞质中或整合到细菌的染色体上。F因子可以在细菌细胞间进行转移并传递遗传物质。第42页,共86页,2023年,2月20日,星期二F因子的存在状态第43页,共86页,2023年,2月20日,星期二接合现象具有F因子的菌株可作为供体,因为F因子中有控制F性伞毛(Fpillus)形成的基因。F性伞毛是由供体细胞表面伸出的一种长附属物,当供体与受体细胞相互接合,F性伞毛就成了两个细胞之间原生质的通道,或叫接合管(conjugationtube)。F+细胞的F因子由接合管向F-传递,使F–受体变成F+。第44页,共86页,2023年,2月20日,星期二F因子及其在杂交中的行为第45页,共86页,2023年,2月20日,星期二F因子及其在杂交中的行为第46页,共86页,2023年,2月20日,星期二F因子及其在杂交中的行为第47页,共86页,2023年,2月20日,星期二F因子及其在杂交中的行为第48页,共86页,2023年,2月20日,星期二(三)、中断杂交试验作图雅各布(Jacob,F)和沃尔曼(Wollman,E.)在五十年代设计了一个著名的中断杂交试验(interruptedmatingexperiment)。他们采用的菌株基因型为:Hfr:thr+leu+aziStonSlac+gal+strSF–:thr–leu–aziRtonRlac–gal-strR这里,thr、leu、lac、gal分别代表苏氨酸、亮氨酸、乳糖和半乳糖,+代表原养型或发酵型,–代表营养缺陷型或不发酵型;azi和tonA分别代表叠氮化纳和T1噬菌体;Str表示链霉素,R代表抗性,S代表敏感。第49页,共86页,2023年,2月20日,星期二中断杂交试验作图将两种细胞混合培养,每隔一定时间取样,把菌液放在食物搅拌器内搅拌,以中断接合。将中断接合的细菌接种到含有链霉素的几种不同培养基上,测定形成了什么样的重组体(链霉素可杀死所有的Hfr供体细胞)。第50页,共86页,2023年,2月20日,星期二中断杂交试验作图结果表明,thr+最先进入F-细胞,接合8分钟后便出现了重组体,leu+随后半分钟出现,aziS在9分钟时出现等等。在被选择的thr+leu+的重组体中,其它的供体基因接连出现,不同的基因经过一定时间就上升到一个稳定的水平。第51页,共86页,2023年,2月20日,星期二中断杂交试验作图第52页,共86页,2023年,2月20日,星期二中断杂交试验作图在检查F-细胞是否得到thr+,可用不加thr而含str、leu的培养基,在这里,只有thr+strR才能生长,能长的细胞就是供体thr+已经进入受体并发生了重组的细胞。试验结果列于表。第53页,共86页,2023年,2月20日,星期二中断杂交试验作图上述事实说明,Hfr菌株的基因是按一定的线性顺序依次进入F–菌株的,染色体从原点以直线方式进入F–细胞。基因位点离原点愈近,进入F–细胞愈早,反之则晚。根据中断杂交的实验,用Hfr基因在F–细胞中出现的时间为标准,可以作出大肠杆菌的遗传连锁图。第54页,共86页,2023年,2月20日,星期二中断杂交试验作图用不同的Hfr菌株进行中断杂交实验所作出的大肠杆菌基因连锁图,其基因向F-细胞转移的顺序大不相同。第55页,共86页,2023年,2月20日,星期二中断杂交试验作图初看起来,似乎每个菌株的基因转移顺序有所不同。其实,转移的顺序并不是随机的。例如,所有的his基因都有gal在一边,gly在另一边。其他基因也是如此,除非它们在连锁群的另一端。这个实验进一步说明F因子和细菌染色体都是环状的,而Hfr细胞染色体的形成,则因F因子插入环状染色体的不同位置,而形成了不同的转移原点和转移方向。
第56页,共86页,2023年,2月20日,星期二中断杂交试验作图第57页,共86页,2023年,2月20日,星期二中断杂交试验作图如果两个基因间的转移时间小于2分钟,用中断杂交法所得的图距不太可靠,应采用传统的重组作图法(recombinationmapping)。例如,有两个紧密连锁的基因:lac+(乳糖发酵)和ade–(腺嘌呤缺陷型),为了求得这两个基因间的距离,可采用Hfrlac+ade+×F–lac–ade–的杂交实验。用完全培养基但不加腺嘌呤,可以选出F–ade+的菌落。由于ade进入F–细胞的顺序较lac为晚,因此,lac–自然也已经进入。如果选出ade+,同时也是lac+,lac–ade间没有发生过交换;如果是lac–,说明两者之间发生过交换。第58页,共86页,2023年,2月20日,星期二基因的重组两基因间的重组频率是22%。但这两个位点间的时间单位约为1分钟,可见1个时间单位(分钟)大约相当于20%的重组值。用重组频率与中断杂交法所测得的基因距离是大致符合的。第59页,共86页,2023年,2月20日,星期二三、性导(sexduction)与F΄因子性导(sexduction)是指接合时由F′因子所携带的外源DNA转移到细菌染色体的过程。F因子整合到宿主细菌染色体的过程是可逆的,当发生环出(loopingout)时,F因子又重新离开染色体。然而F因子偶然在环出时不够准确,它携带有染色体的一些基因。阿代尔伯格和伯恩斯(Adelberg,E.和Burns,S.,1959)称这种F因子为F′因子。第60页,共86页,2023年,2月20日,星期二性导(sexduction)与F΄因子F′因子使细菌带有某些突出的特点:第一,F′因子以极高的比率转移它的基因,如同F+细菌以极高的比率转移它的F+因子一样;第二,F′因子有极高的自然整合率,而且整合在一定的座位上,因为它有与细菌染色体的同源区段。
第61页,共86页,2023年,2月20日,星期二性导(sexduction)与F΄因子性导在大肠杆菌的遗传学研究中十分有用。第一,不同的F′因子带有不同的细菌DNA片段,利用不同的F′的性导可以测定不同基因在一起转移的频率。其次,观察由性导形成的杂合部分二倍体中某一性状的表现,可以确定这一性状的等位基因的显隐关系。第三,性导形成的部分二倍体也可用作互补测验,确定两个突变型是同属于一个基因还是不同基因。第62页,共86页,2023年,2月20日,星期二第三节噬菌体的遗传分析一、烈性噬菌体二、温和噬菌体三、转导第63页,共86页,2023年,2月20日,星期二噬菌体的遗传分析
遗传学上应用最广泛的是大肠杆菌的T噬菌体系列(T1到T7)。T偶列噬菌体具有六角形的头部,其内含有双链DNA分子。头下的尾部包括一个中空的针状结构及外鞘。末端是基板,由尾丝及尾针组成。通过尾鞘的收缩将噬菌体DNA经中空尾部注入宿主细胞。根据噬菌体DNA在宿主细菌内的特点,又将噬菌体分为两类。第64页,共86页,2023年,2月20日,星期二一、烈性噬菌体噬菌体的遗传物质经中空尾部进入宿主细胞,遂即破坏宿主细胞原有的遗传物质,并转而合成大量的噬菌体遗传物质和蛋白质,组装成许多新的子噬菌体,最后使细菌裂解(lysis)。第65页,共86页,2023年,2月20日,星期二二、温和噬菌体温和性噬菌体具有溶源性(lysogeny)的生活周期,即在噬菌体侵入后,细菌并不裂解,它们以两种不同形式出现,λ和P1噬菌体各代表一种略有不同的溶原性类型。λ噬菌体附着于大肠杆菌染色体的gal和bio位点之间的attλ座位上(attachmentsite),它能通过交换而整合到细菌染色体上。这时它会阻止其他λ噬菌体的超数感染(superinfection)(一个细菌受一个以上噬菌体所感染的现象)。整合的噬菌体称为原噬菌体(prophage)。P1噬菌体与λ不同,它并不整合到细菌的染色体上,而是独立地存在于它的细胞质内。第66页,共86页,2023年,2月20日,星期二温和噬菌体第67页,共86页,2023年,2月20日,星期二温和噬菌体这两种噬菌体的共同特点是核酸既不大量的复制,也不大量的转录和翻译。这类噬菌体往往只有一个或少数基因表达,由此产生的阻遏物能关闭其他基因的表达。当溶源性细菌分裂成两个子细胞时,噬菌体λ随细菌染色体的复制而复制,每个子细胞中有一个拷贝。噬菌体P1的复制则使每个子细胞中至少含有一个拷贝。原噬菌体通过诱导(induction)可转变为烈性噬菌体。诱导可以通过不同的方式进行,如UV照射、温度改变、与非溶源性细菌的接合等。这类诱导可以造成阻遏物失活或稀释,使其他的噬菌体基因表达出来,促使噬菌体繁殖并进入裂解周期。第68页,共86页,2023年,2月20日,星期二三、转导(transduction)
转导(transduction)是指以噬菌体为媒介所进行的细菌遗传物质重组的过程。它与上述的转化、性导、接合的主要不同之处,在于它是以噬菌体为媒介的。在这一过程中,细菌的一段染色体被错误地包装在噬菌体的蛋白质外壳内,并通过感染而转移到另一个受体细菌内。第69页,共86页,2023年,2月20日,星期二转导(transduction)黎德伯格等在鼠伤寒沙门氏菌中发现转导现象。用两个营养缺陷型杂交,一个不能合成苯丙氨酸、色氨酸和酪氨酸(phe-、trp-、tryr-),另一个不能合成甲硫氨酸和组氨酸(met-、his-),结果在基本培养基上发现原养型的菌落。为了确定是否发生了接合,将两种菌株分别放在U型管的两臂内,中间用玻璃滤板隔开,以防止细胞直接接触,但可以允许比细菌小的物质通过,竟然也获得了野生型重组型。第70页,共86页,2023年,2月20日,星期二转导(transduction)可能的结论是,这种重组是通过一种过滤性因子(filterableagent,FA)而实现的。由于FA不受DNA酶的影响,这样就消除了转化作用的可能性。进一步研究证明,FA是一种噬菌体,称为P22,它对亲本菌株之一是溶源性的。支持这个结论的证据是:(1)FA的大小和质量与P22相同;(2)FA用抗P22血清处理后失活。转导可分为普遍性转导(generalizedtransduction)和特殊性转导(specializedtransduction)两大类。第71页,共86页,2023年,2月20日,星期二转导(transduction)第72页,共86页,2023年,2月20日,星期二普遍性转导
1.普遍性转导在噬菌体感染的末期,细菌染色体被断裂成许多小片段,在形成噬菌体颗粒时,少数噬菌体将细菌的DNA误认做是它们自己的DNA,并以其外壳蛋白将其包围,从而形成转导噬菌体。由于可以转导细菌染色体组的任何不同部分,因此称为普遍性转导。这种假噬菌体称为转导颗粒(transducingparticle)。这种颗粒既然不携带噬菌体基因,对受体细菌就没有有害的影响。当转导颗粒将它的内含物注入受体细菌后,形成一个部分二倍体,导入的基因经过重组,整合到宿主的染色体上。由此形成的具有重组遗传结构的细菌细胞叫做转导体(transductant)。第73页,共86页,2023年,2月20日,星期二普遍性转导两个基因同在一起转导称为合转导(cotransduction),合转导的频率愈高,表明两个基因在染色体上的距离愈近,连锁愈密切;相反,如果两个基因的合转导频率很低,就说明它们之间距离较远。通过观察两因子转导(two-factortransduction),计算并比较每两个基因之间的合转导频率,就可以确定三个基因或三个以上基因在染色体上的排列顺序。例如a基因和b基因的合转导频率很高,和c基因的合转导频率也很高,而b和c很少或完全不在一起转导,这三个基因的次序就应为bac。如果研究三因子转导(three-factortransduction),只需分析一个实验的结果就可以推出三个基因的次序。第74页,共86页,2023年,2月20日,星期二普遍性转导
例如,供体大肠杆菌具有基因型a+b+c+,受体的基因型为a-b-c-。供体用P1噬菌体感染,P1的后代再用来感染受体细胞,然后把受体细胞接种在选择培养基上。如果通过中断杂交已知三个基因中的一个如a不在中间,就可对a+进行选择,即在对a+进行选择的选择培养基上,把可以生长的a+细胞选出来。然后,再把被选择的受体细胞重复接种在其他对b+或c+进行选择的选择培养基上,检查a+细胞是否同时具有b+和c+。最少的一类转导体应当代表最难于转导的情况,这种转导体是同时发生交换次数最多的一类。它的两边应为供体基因,而中间为受体基因,如正确次序为abc,就应为a+b-c+。假定由实验得到的最少的转导体类别为a+b+c-,那么就可以确定,这三个基因的正确次序应当是acb或bca,而不是abc。第75页,共86页,2023年,2月20日,星期二普遍性转导第76页,共86页,2023年,2月20日,星期二普遍性转导如果把三个基因中每两个基因的合转导频率算出来,还可推算出三个基因之间的物理距离。若两个基因紧密连锁,就可能经常在一起转导,合转导频率将接近于1。如果两个基因从来或者几乎不包含在同一转导DNA片段中,它们的合转导频率接近于或等于0。利用这种关系可以求出同一染色体上两个基因之间的物理距离。经推导,得到以下计算公式:d=同一染色体上两基因之间的物理距离。L=转导DNA的平均长度。.X=两个基因合转导的频率。转导颗粒DNA的平均长度(约为1个病毒基因组的大小)知道后,就可以依上述公式估算出两个较近基因间的分子距离。第77页,共86页,2023年,2月20日,星期二普遍性转导
p1噬菌体可以携带大肠杆菌DNA的任何部分,在分析大肠杆菌基因的连锁关系上是很有效的。例如,先利用普遍性转导噬菌体P1,侵染带有leu+、thr+、aziR三个基因的大肠杆菌,再用来自后者(供体)的P1侵染带有leu-、thr-、aziS
的三个标记基因的大肠杆菌(受体),然后将受体细菌进行特定培养,以测定该三个基因的连锁关系。其方法是把受体细菌培养在一种可以选择1-2个标记基因而不选择其余标记基因的培养基上。例如,把受体细菌放在没有叠氮化钠(azi)但加有苏氨酸(thr)的基本培养基上培养,于是leu就成为选择的标记基因,因为在该培养基上只有leu+细胞才能生长。thr和azi是未选择的标记基因,因为当培养基内有thr时,其标记基因可能是thr+或thr–;当培养基内无叠氮化钠时,其标记基因可能是aziR或aziS。对每个选择的标记基因进行多次实验,以
温馨提示
- 1. 本站所有资源如无特殊说明,都需要本地电脑安装OFFICE2007和PDF阅读器。图纸软件为CAD,CAXA,PROE,UG,SolidWorks等.压缩文件请下载最新的WinRAR软件解压。
- 2. 本站的文档不包含任何第三方提供的附件图纸等,如果需要附件,请联系上传者。文件的所有权益归上传用户所有。
- 3. 本站RAR压缩包中若带图纸,网页内容里面会有图纸预览,若没有图纸预览就没有图纸。
- 4. 未经权益所有人同意不得将文件中的内容挪作商业或盈利用途。
- 5. 人人文库网仅提供信息存储空间,仅对用户上传内容的表现方式做保护处理,对用户上传分享的文档内容本身不做任何修改或编辑,并不能对任何下载内容负责。
- 6. 下载文件中如有侵权或不适当内容,请与我们联系,我们立即纠正。
- 7. 本站不保证下载资源的准确性、安全性和完整性, 同时也不承担用户因使用这些下载资源对自己和他人造成任何形式的伤害或损失。
最新文档
- 西安市新城区2025年四下数学期末考试试题含解析
- 苏州市职业大学《制药技术与设备》2023-2024学年第二学期期末试卷
- 佳木斯市2024-2025学年五下数学期末监测试题含答案
- 江西理工大学《中华传统文化》2023-2024学年第二学期期末试卷
- 河海大学《第二外语日(一)》2023-2024学年第一学期期末试卷
- 江西省上饶市四中重点中学2024-2025学年初三下学期期末质检生物试题试卷含解析
- 浙江省镇海中学2025年新高三下开学适应性考试生物试题试卷含解析
- 西藏林芝地区朗县2024-2025学年四年级数学第二学期期末监测模拟试题含解析
- 昆明卫生职业学院《社会化营销案例研究》2023-2024学年第二学期期末试卷
- 枣庄学院《中国花鸟画工笔》2023-2024学年第二学期期末试卷
- 饲料学全套课件
- 血液透析相关无菌技术操作护理课件
- 注塑产品常见外观缺陷课件
- 未来人工智能在麻醉学中的应用前景分析培训课件
- 医院培训课件:《枸橼酸钠抗凝剂在CRRT中的应用》
- 戊二酸血症 I 型学习课件
- 发展全过程人民民主发展全过程人民民主
- 烟花爆竹零售经营安全培训
- (2023年度)中央厨房、集体供餐配送单位食品安全全项目自查记录表
- 猕猴桃果酱制作方法
- 逆变器行业营销策略方案
评论
0/150
提交评论