



版权说明:本文档由用户提供并上传,收益归属内容提供方,若内容存在侵权,请进行举报或认领
文档简介
1、1983. Barbara McClintock DNA transposable elememtIS-两端有 IR,只编码转座酶类转座因子-结构同 IS,但不能独立存在,仅作为复合转座子的两端组件复合转座子-两端由 IS 或类 IS 构成,可编码抗抗菌素物质原核TnA 转座子家族-两端为 IR,可编码转座酶、解离酶和抗性物质AC-Ds-植物(玉米)中的激活-解离因子转座子P 因子-果蝇中父本因子,在 MP中导致杂种不育反转录病毒、RNA DNA整合宿主靶 DNATyCopia病毒超家族LINSL1(1)有长末端重复序列(2)编码反转录酶或整合酶(3)可含内含子SINSB1/ Alu(1)无重
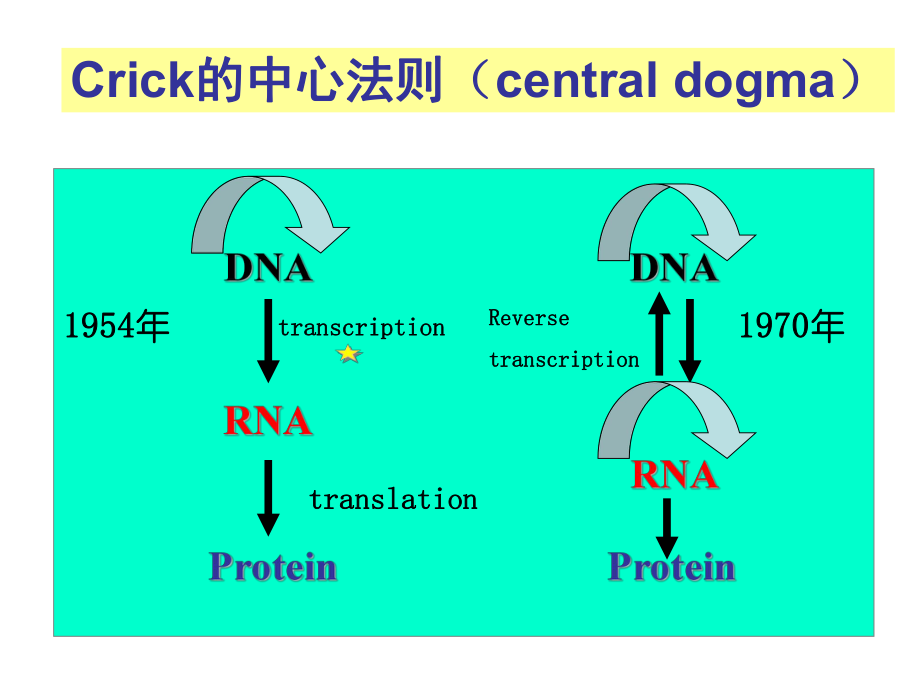
2、复序列(2)不编码转座子产物(3)无内含子转座因子真核反转录转座子非病毒超家族假基因第三讲第三讲 生物信息的传递生物信息的传递(上)从(上)从DNA到到RNA Crick的中心法则的中心法则(central dogma)19541954年年transcriptiontranscriptiontranslationtranslation19701970年年ReverseReversetranscriptiontranscriptionFrom DNA to Protein DNA序列是遗传信息序列是遗传信息的贮存者,它通过自的贮存者,它通过自主复制得到永存,并主复制得到永存,并通过通过转录转录生
3、成生成信使信使RNA,翻译翻译生成生成蛋白蛋白质质的过程来控制生命的过程来控制生命现象。现象。基因表达包括基因表达包括转录转录(transcription)和)和翻译翻译(translation)两个阶段。)两个阶段。转录转录是指拷贝出一条与是指拷贝出一条与DNA链序列完全相链序列完全相同同(除了除了TU之外)的之外)的RNA单链的过程,是单链的过程,是基因表达的核心步骤基因表达的核心步骤。翻译翻译是指以新生的是指以新生的mRNA为模板,把核苷酸为模板,把核苷酸三联遗传密码子翻译成氨基酸序列、合成多三联遗传密码子翻译成氨基酸序列、合成多肽链的过程,是肽链的过程,是基因表达的最终目的基因表达的最
4、终目的。DNA-mRNA-the encoded peptide (sense strand)(antisense strand)编码链(编码链(coding strand): 与与mRNA序列相同的那条序列相同的那条DNA链链, 或或称有称有意义链(意义链(sense strand)。)。模板链(模板链(template strand): 根据碱基互补原则指导根据碱基互补原则指导mRNA合合成的成的DNA链链, 或称或称反义链(反义链(antisense strand)。)。35new RNA strand3535template strand*coding strand*GTACGUACC
5、ATGnegative strandantisense strandpositive strandsense strandWatson strandCrick strandRNA主要以单链形式存在于生物体内主要以单链形式存在于生物体内生物体内拥有生物体内拥有三类主要三类主要RNARNA:1、编码特定蛋白质序列的、编码特定蛋白质序列的mRNA;2、能特异性解读、能特异性解读mRNA 中的遗传信息并中的遗传信息并将其转化成相应氨基酸后加入多肽链中将其转化成相应氨基酸后加入多肽链中的的tRNA;3、直接参与核糖体中蛋白质合成的、直接参与核糖体中蛋白质合成的rRNA。RNA的转录的转录(Transcr
6、iption) RNA 是以是以53方向方向合成的,它的序列与合成的,它的序列与DNA编码链(意义链)相同。编码链(意义链)相同。 RNA 的合成是的合成是以反义链以反义链(模板链模板链)为模板为模板。 同在同在DNA中一样,中一样,形成磷酸二酯键形成磷酸二酯键( Phosphodiester bonds)。 必需的成分必需的成分: RNA 聚合酶聚合酶, rNTPs, 转录调控因子转录调控因子, 启动启动子子 & 终止子终止子/模板模板RNA合成的特点合成的特点3.1 转录的基本过程转录的基本过程 1、模板识别、模板识别 (Template Recognition) 2、转录起始、转
7、录起始 (Initiation) 3、转录的延伸、转录的延伸 (Elongation)4、转录的终止、转录的终止 (Termination)1、RNA聚合酶与启动子聚合酶与启动子DNA双链相互双链相互作用并与之相结合的过程。作用并与之相结合的过程。模板识别模板识别2、转录起始前,启动子附近的、转录起始前,启动子附近的DNA双双链分开形成链分开形成转录泡转录泡以促使底物核糖核苷以促使底物核糖核苷酸与模板酸与模板DNA的碱基配对。的碱基配对。1.在起始位点合成在起始位点合成RNA链:第一个核苷酸链:第一个核苷酸键的产生,该位点被称为键的产生,该位点被称为 +1 。转录起始转录起始2. 转录起始后直
8、到形成转录起始后直到形成9个核苷酸短链是通个核苷酸短链是通过启动子阶段。过启动子阶段。3. 通过启动子的时间代表一个启动子的强通过启动子的时间代表一个启动子的强弱弱:时间越短时间越短,转录起始的频率也越高转录起始的频率也越高。转录起始转录起始就是就是RNA链上链上第一个核苷第一个核苷酸键的产生。酸键的产生。 一般情况下,转录起始复合物可以进入两一般情况下,转录起始复合物可以进入两条不同的反应途径:条不同的反应途径:合成并释放合成并释放2-9个核苷酸的短个核苷酸的短RNA转录物,即转录物,即所谓的所谓的流产式起始流产式起始;一旦一旦RNA聚合酶成功地合成聚合酶成功地合成9个以上核苷酸,个以上核苷
9、酸,尽快释放尽快释放亚基,亚基,RNA聚合酶离开启动子区,聚合酶离开启动子区,转录就进入正常的延伸阶段。转录起始复合物转录就进入正常的延伸阶段。转录起始复合物通过上游启动子区并生成由核心酶、通过上游启动子区并生成由核心酶、DNA和新和新生生RNA所组成的所组成的转录延伸复合物转录延伸复合物。封闭复合物封闭复合物(closed complex)开放复合物开放复合物(open complex)转录起始包括三步转录起始包括三步三元复合物三元复合物(ternary complex )封闭复合物(封闭复合物(closed complex)聚合酶与启动子可逆性结合形成封闭复合物。聚合酶与启动子可逆性结合形
10、成封闭复合物。 DNA 链仍处于双链状态。链仍处于双链状态。19开放复合物(开放复合物(open complex) 聚合酶结合的聚合酶结合的DNA序列中有一小段双链序列中有一小段双链被解开,被解开, 14 bp (-11 to +3 ) around the start site (+1 site) 转录泡形成转录泡形成 三元复合物三元复合物(ternary complex ) 开放复合物与最初的两个开放复合物与最初的两个NTP相结合并在这两相结合并在这两个核苷酸之间形成磷酸二酯键。个核苷酸之间形成磷酸二酯键。 三元复合物三元复合物=DNA +RNA + enzymeRNA合成的起始合成的起始
11、 RNA聚合酶全酶聚合酶全酶+启动子启动子DNA处于双链状态处于双链状态聚合酶全酶所结合的聚合酶全酶所结合的DNA序序列中有一小段双链被解开列中有一小段双链被解开RNA聚合酶、聚合酶、DNA和新生和新生RNA 转录的延伸转录的延伸RNA聚合酶离开启动聚合酶离开启动子,沿子,沿DNA链移动并使链移动并使新生新生RNA链不断伸长的链不断伸长的过程就是过程就是转录的延伸转录的延伸。 共价地向生长共价地向生长RNA链的链的3端添加核糖核苷酸端添加核糖核苷酸 RNA聚合酶是以聚合酶是以5 3 方向来延长方向来延长RNA链链 RNA聚合酶本身沿着反义链以聚合酶本身沿着反义链以3 5方向移动方向移动 大肠杆
12、菌大肠杆菌RNA聚合酶的活性一般为每秒聚合酶的活性一般为每秒40-90个核苷酸。个核苷酸。随着随着RNA聚合酶的移动,聚合酶的移动,DNA双螺旋持续双螺旋持续解开,暴露出新的单链解开,暴露出新的单链DNA模板模板, 新生新生RNA链的链的3末端不断延伸,在解链区形成末端不断延伸,在解链区形成RNA-DNA杂合物。杂合物。转录的终止转录的终止 转录的终止:转录的终止: 当当RNA链延伸到转录终止位点时,链延伸到转录终止位点时,RNA polymerase 和和RNA链均从链均从DNA模板上释放出模板上释放出来。来。 Terminator: 通常含有自我互补区域通常含有自我互补区域(self-co
13、mplementary regions) ,在,在RNA产物中可以形成产物中可以形成stem-loop 或或hairpin结构。结构。转录生成转录生成mRNA的速度大约是每分钟的速度大约是每分钟2500个核苷酸个核苷酸(14个密码子个密码子/秒秒),与翻译,与翻译速度速度(15aa/秒秒)基本相等,但比基本相等,但比DNA复制复制的速度要慢得多。的速度要慢得多。常识数据常识数据基因开始表达基因开始表达mRNA的间隔约为的间隔约为2.5分分钟,而再过半分钟就能在细胞内测到相钟,而再过半分钟就能在细胞内测到相应的蛋白质。应的蛋白质。Two types of terminators in E. co
14、li不依赖于不依赖于因子的终止因子的终止 intrinsic terminator (内在终止子内在终止子) 依赖于依赖于因子的终止因子的终止不依赖于不依赖于因子的终止因子的终止由两个序列元件组成:由两个序列元件组成:一段短的反向重复序列一段短的反向重复序列(大约(大约20个核苷酸)个核苷酸);其后是一段大约其后是一段大约4-8个个A:T碱基对的序列碱基对的序列。由它转录出由它转录出mRNA可形可形成发夹结构,可阻止成发夹结构,可阻止RNA pol的前进的前进。不牢固的碱基对不牢固的碱基对: A:U 容易断裂容易断裂不依赖于不依赖于因子因子的终止子的终止子 (内在内在终止子终止子 ) 终止效率
15、与二重对终止效率与二重对称序列和寡聚称序列和寡聚U U的的长短有关,长度长短有关,长度 效率效率 依赖于依赖于因子的终止因子的终止提纯的提纯的RNARNA聚合酶并不能识别特异性的转录终聚合酶并不能识别特异性的转录终止信号,而加入止信号,而加入大肠杆菌大肠杆菌因子因子后该聚合酶就后该聚合酶就能在能在DNADNA模板上准确地终止转录。模板上准确地终止转录。只含有自我互补区域,只含有自我互补区域,可形成茎环结构,但在茎中的可形成茎环结构,但在茎中的G.C含量少,茎环易打开。含量少,茎环易打开。 其终止需要其终止需要 因子的参与。因子的参与。 因子与因子与ssRNA的特定位点结合的特定位点结合(C丰富
16、,丰富,G缺乏缺乏 )。通过催化通过催化NTP的水解促使新生的水解促使新生RNA链从三元转录复链从三元转录复合物中解离。合物中解离。因子因子因子因子是一个相对分子是一个相对分子质量为质量为2.0105的的六聚六聚体体蛋白,可水解各种蛋白,可水解各种核苷三磷酸促使新生核苷三磷酸促使新生RNA链从三元转录复链从三元转录复合物中解离出来,从合物中解离出来,从而终止转录。而终止转录。结合到结合到RNA链终止子上游的链终止子上游的某一序列某一序列 因子结合以后延着因子结合以后延着RNA向向3端移动,跟踪聚合酶端移动,跟踪聚合酶 追上在终止位点暂停的追上在终止位点暂停的RNA 聚合酶聚合酶终止终止-三元复
17、合物解体三元复合物解体因子参与的因子参与的RNA合成终止模式合成终止模式“穷追穷追”(hot pursuit)模型)模型 转录机器的主要成分转录机器的主要成分 RNA聚合酶聚合酶 (RNA polymerase) 转录复合物转录复合物 RNA聚合酶聚合酶 (RNA polymerase)RNA聚合酶聚合酶是转录过程中是转录过程中最关键最关键的酶的酶。主要以双链主要以双链DNA为模板,以为模板,以4种核苷三磷酸种核苷三磷酸作为活性前体。作为活性前体。需要需要Mg2+/Mn2+为辅助因子。为辅助因子。它不需要任何引物它不需要任何引物。以以5 3 方向合成方向合成RNA链。链。缺乏缺乏3 5 外切酶
18、活性。外切酶活性。1. 是一个含有多个亚单位是一个含有多个亚单位(multi-subunit)的酶。的酶。识别识别DNA双链上的启动子;双链上的启动子;(2) 使使DNA变性,在启动子处解旋变性,在启动子处解旋成单链;成单链;(3) 通过阅读启动子序列,通过阅读启动子序列,RNA pol 确定它自己的转录方向和模板确定它自己的转录方向和模板链。链。(4) 最后当它达到终止子时,通过最后当它达到终止子时,通过识别停止转录。识别停止转录。 RNA聚合酶需执行的功能聚合酶需执行的功能原核原核和和真核真核生物的生物的RNA聚合酶虽然聚合酶虽然都能催化都能催化RNA的合成,但在其分子的合成,但在其分子组
19、成、种类和生化特性上各有特色。组成、种类和生化特性上各有特色。2 alpha () subunit,1 beta () subunit, 1 beta prime () subunit, 1 omega () subunit,1 sigma () subunit原核生物原核生物(E.coli)的的RNA聚合酶聚合酶Core Enzyme核心酶核心酶Holoenzyme聚合酶全酶聚合酶全酶相对分子量:相对分子量:4.65105参与转录延伸参与转录延伸只与转录的只与转录的起始有关起始有关36.5 KD36.5 KD151 KD155 KD11 KD70 KD原核生物原核生物(E.coli)的的RN
20、A聚合酶聚合酶 E. coli 只有只有一个一个 DNA指导的指导的 RNA聚合酶聚合酶 ,来,来合成所有类型的合成所有类型的RNA。 是细胞中最大的酶之一。是细胞中最大的酶之一。 由由5种亚基组成聚合酶全酶种亚基组成聚合酶全酶(holoenzyme),包括包括 2 , 1 , 1, 1 以及以及1 亚基。亚基。 形状象一个圆筒状通道,可以直接与形状象一个圆筒状通道,可以直接与16 bp DNA结合。整个聚合酶可结合结合。整个聚合酶可结合40 bp DNA。 RNA 合成速率合成速率: 40 nt /秒秒, 37oC 。是核心酶中的两个相同的亚单位是核心酶中的两个相同的亚单位由由rpoA 基因
21、编码基因编码与核心酶的组装及启动子识别有关与核心酶的组装及启动子识别有关参与参与RNA聚合酶和部分调节因子的相互作用聚合酶和部分调节因子的相互作用E. coli RNA polymerase :subunit 和和分别由分别由 rpoB 和和 rpoC 基因编码。基因编码。E. coli RNA polymerase : & subunit由由和和亚基亚基组成了聚合酶的催化中心组成了聚合酶的催化中心。它们在序列上与真核生物它们在序列上与真核生物RNA聚合酶的两聚合酶的两个大亚基有同源性。个大亚基有同源性。亚基能与模板亚基能与模板DNA、新生、新生RNA链及核苷链及核苷酸底物相结合。酸底
22、物相结合。 rpoB 和和 rpoC 基因的突变基因的突变会影响转录所有的阶段。会影响转录所有的阶段。负责负责模板链的选择和转录的起始模板链的选择和转录的起始:E. coli RNA polymerase : factor与与因子的结合使因子的结合使RNA聚合酶从核心酶转变为聚聚合酶从核心酶转变为聚合酶全酶。合酶全酶。是是启动子识别的关键的酶启动子识别的关键的酶,不仅增加聚合酶对启,不仅增加聚合酶对启动子的亲和力动子的亲和力(提高提高103倍倍),还可降低它对非特异,还可降低它对非特异性位点的亲和力性位点的亲和力(降低降低104倍倍),使酶底复合物的半,使酶底复合物的半衰期小于衰期小于1s。在
23、细胞中对在细胞中对因子量的需求少于聚合酶中其它亚因子量的需求少于聚合酶中其它亚单位。单位。在某些细菌中含有识别不同启动子在某些细菌中含有识别不同启动子的的因子,以适应不同生长发育阶因子,以适应不同生长发育阶段的要求,控制不同基因转录的起段的要求,控制不同基因转录的起始。如枯草芽孢杆菌中始。如枯草芽孢杆菌中5555主要在主要在营养细胞中,营养细胞中,2929则主要出现在孢则主要出现在孢子形成阶段,参与孢子形成期基因子形成阶段,参与孢子形成期基因转录的调控。转录的调控。 T3 和和 T7 噬菌体的噬菌体的RNA 聚合酶是由一条小的聚合酶是由一条小的多肽链组成多肽链组成, 相对分子量小于相对分子量小
24、于1105; 在在37C下,其转录速度为下,其转录速度为 200 nt/秒;秒; 它们只能识别不同于它们只能识别不同于E. coli 启动子的自身的启动子的自身的启动子。启动子。RNA polymerase differs from organism to organism有有3类类RNA聚合酶聚合酶;真核生物的真核生物的RNA聚合酶聚合酶结构比大肠杆菌结构比大肠杆菌RNA聚合酶复杂;聚合酶复杂;在细胞核中的位置不同;在细胞核中的位置不同;负责转录的基因不同,对负责转录的基因不同,对-鹅膏蕈碱的敏感鹅膏蕈碱的敏感性也不同。性也不同。真核生物真核生物RNA聚合酶一般有聚合酶一般有8-16个亚基所
25、组成,个亚基所组成,相对分子质量超过相对分子质量超过5105。真核细胞中三类真核细胞中三类RNA聚合酶特性比较聚合酶特性比较酶酶细胞内细胞内定位定位转录产转录产物物相对活相对活性性对对-鹅膏鹅膏蕈碱的敏蕈碱的敏感程度感程度RNA聚合聚合酶酶I核仁核仁rRNA(28S,18S,5.8S)50%-70%不敏感不敏感RNA聚合聚合酶酶II核质核质hnRNA*,snRNA,mRNA20%-40%敏感敏感RNA聚合聚合酶酶III核质核质tRNA5SrRNA某些涉及某些涉及RNA加工的加工的snRNA约约10%存在物种存在物种特异性特异性*hnRNA: heterogeneous muclear RNA,
26、核内不均一RNA,mRNA的前体在动、植物及昆虫的细胞中,在动、植物及昆虫的细胞中,RNA RNA pol pol 的活性可被低浓度的的活性可被低浓度的-鹅膏鹅膏蕈蕈碱所抑制。但却不抑制碱所抑制。但却不抑制pol pol 。Pol Pol 对对-鹅膏鹅膏蕈蕈碱的反应,不同碱的反应,不同的生物有所差异。在动物细胞中高浓的生物有所差异。在动物细胞中高浓度的度的-鹅膏鹅膏蕈蕈可抑制转录,在昆虫可抑制转录,在昆虫中不受抑制。中不受抑制。转录的抑制剂转录的抑制剂抑制剂抑制剂靶酶靶酶抑制作用抑制作用利福霉素利福霉素细菌全酶细菌全酶和和亚基结合,抑亚基结合,抑制起始制起始链霉溶菌素链霉溶菌素细菌核心酶细菌核
27、心酶 和和亚基结合,抑亚基结合,抑制起始制起始放射线素放射线素D真核真核Pol和和DNA结合,阻止结合,阻止延伸延伸-鹅膏蕈碱鹅膏蕈碱真核真核Pol和和RNA Pol结合结合RNA合成抑制剂主要分两类合成抑制剂主要分两类:1. 模板结合抑制剂模板结合抑制剂2. 聚合酶抑制剂聚合酶抑制剂聚合酶中有两个相对分子质量超聚合酶中有两个相对分子质量超过过1105的大亚基;的大亚基;真核生物真核生物RNA聚合酶的主要特征聚合酶的主要特征同种生物同种生物3类聚合酶有类聚合酶有“共享共享”小小亚基的倾向,即有几个小亚基是其亚基的倾向,即有几个小亚基是其中中3类或类或2类聚合酶所共有的。类聚合酶所共有的。真核生
28、物真核生物RNA聚合酶的亚基聚合酶的亚基RNARNA聚合酶聚合酶I IRNARNA聚合酶聚合酶IIIIRNARNA聚合酶聚合酶IIIIIIRPA 1RPA 1RPB 1 (RPB 1 ( ) )RPC 1RPC 1RPA 2RPA 2RPB 2 (RPB 2 ( ) )RPC 2RPC 2RPC 5RPC 5RPB 3(RPB 3() )RPC 5RPC 5RPC 9RPC 9RPB 11 (RPB 11 () )RPC 9RPC 9RPB 6RPB 6RPB 6 (RPB 6 () )RPB 6RPB 6其它其它9 9个亚基个亚基其它其它7 7个亚基个亚基其它其它1111个亚基个亚基注:亚基
29、按照分子量由大到小的顺序排列。真核生物真核生物RNA聚合酶一般有聚合酶一般有8-16个亚基所组成个亚基所组成, 相对分相对分子质量超过子质量超过5105。真核生物线粒体和叶绿体中存在真核生物线粒体和叶绿体中存在不同的不同的RNA聚合酶聚合酶线粒体中线粒体中RNA聚合酶:聚合酶: 只有一条多肽链,相对分子量小于只有一条多肽链,相对分子量小于7 X 104,是已知,是已知最小的最小的RNA聚合酶之一;聚合酶之一; 与与T7噬菌体噬菌体RNA聚合酶有同源性。聚合酶有同源性。叶绿体中叶绿体中RNA聚合酶:聚合酶: 比较大;比较大; 结构上与细菌中的聚合酶相似,由多个亚基组成,结构上与细菌中的聚合酶相似
30、,由多个亚基组成,部分亚基由叶绿体基因编码。部分亚基由叶绿体基因编码。线粒体和叶绿体线粒体和叶绿体RNA聚合酶活性不受聚合酶活性不受-鹅膏蕈鹅膏蕈碱所抑制。碱所抑制。RNA polymerase/transcription and DNA polymerase/replicationRNA polDNA pol模板模板dsDNAdsDNA需要引物需要引物NoYes启动启动promoterorigin延伸速度延伸速度40 nt/ sec900 bp/sec核酸外切酶活性核酸外切酶活性 NoYes终止子终止子Synthesized RNA Template DNA转录机器的主要成分转录机器的主要成
31、分 RNA聚合酶聚合酶 (RNA polymerase) 转录复合物转录复合物 封闭复合物封闭复合物开放复合物开放复合物三元复合物三元复合物RNA合成起始合成起始RNA聚合酶全酶聚合酶全酶+启动子启动子DNA处于双链状态处于双链状态聚合酶全酶所结合的聚合酶全酶所结合的DNA序序列中有一小段双链被解开列中有一小段双链被解开RNA聚合酶、聚合酶、DNA和新生和新生RNA转录复合物转录复合物 除除RNA聚合酶之外,聚合酶之外,真核生物真核生物转转录起始过程中至少还需要录起始过程中至少还需要7种辅种辅助因子参与,这些蛋白辅助因子助因子参与,这些蛋白辅助因子统称为转录因子(统称为转录因子(transcr
32、iption factor, TF)。)。真核生物真核生物RNA聚合酶聚合酶II所形成的转录起始复合物所形成的转录起始复合物蛋白质蛋白质亚基数亚基数亚基的分子量亚基的分子量(X103)功功 能能RNA聚合酶聚合酶II1210-220催化催化RNA的生物合成的生物合成TBP 138与启动子上的与启动子上的TATA区相结合区相结合TF II A312,19,35使使TBP及及TFIIB与启动子的结合比与启动子的结合比较稳定较稳定TF II B135与与TBP相结合,吸引相结合,吸引RNA聚合酶和聚合酶和TFIIF到启动区上到启动区上TF II D1215-250与各种调控因子相互作用与各种调控因子
33、相互作用TF II E234,57吸引吸引TFIIH,有,有ATP酶及解链酶活酶及解链酶活性性TF II F230,74结合结合RNA聚合酶聚合酶II并在并在TFIIB帮助帮助下阻止聚合酶与非特异性下阻止聚合酶与非特异性DNA序列序列相结合相结合TF II H1235-89在启动子区解开在启动子区解开DNA双链,使双链,使RNA聚合酶聚合酶II磷酸化,接纳核苷酸磷酸化,接纳核苷酸切除修复体系切除修复体系真核生物真核生物RNA聚合酶不能直接识别基因聚合酶不能直接识别基因的启动子区,需要一些被称为的启动子区,需要一些被称为转录调控转录调控因子因子的辅助蛋白质按特定顺序结合于启的辅助蛋白质按特定顺序
34、结合于启动子上,动子上,RNA聚合酶才能与之相结合并聚合酶才能与之相结合并形成复杂的形成复杂的前起始复合物前起始复合物(PIC),以保证以保证有效地起始转录。有效地起始转录。前起始复合物前起始复合物(preinitiation transcription complex, PIC)转录起点转录起点是指与新生是指与新生RNA链第一个核苷酸相对应链第一个核苷酸相对应DNA链上的碱基,通常为链上的碱基,通常为嘌呤嘌呤。把起点把起点55末端末端的序列称为的序列称为上游(上游(upstreamupstream),),把其把其33末端末端的序列称为的序列称为下游(下游(downstreamdownstre
35、am)。)。转录单元(转录单元(transcription unit) 在原核中在原核中90%为嘌呤,为嘌呤,A或或G; 转录起始位点转录起始位点 (Transcription start site) 位置固定位置固定 ; 通常在起始核苷酸的两侧为通常在起始核苷酸的两侧为C 和和 T (i.e. CGT or CAT) 启动子区的基本结构启动子区的基本结构启动子启动子是一段位于结构基因是一段位于结构基因55端上游端上游区区的保守的的保守的DNADNA序列,能序列,能活化活化RNARNA聚合酶聚合酶,使之与模板使之与模板DNADNA准确地相结合并具有准确地相结合并具有转转录起始的特异性录起始的特
36、异性。启动子与转录起始启动子与转录起始大肠杆菌大肠杆菌RNA聚合酶与启动子的相互作用主聚合酶与启动子的相互作用主要包括:要包括: 启动子区的识别;启动子区的识别; 酶与启动子的结合;酶与启动子的结合; 因子的结合与解离。因子的结合与解离。原核生物不同基因的启动子原核生物不同基因的启动子1975年,年,Pribnow和和Schaller将将RNA聚合酶全酶与模板聚合酶全酶与模板DNA结合后,用结合后,用DNaseI降解降解DNA,得到,得到4144个核个核苷酸对的苷酸对的DNA片段。片段。10区区(Pribnow区区)的发现的发现序列分析发现,在被保护区内有一序列分析发现,在被保护区内有一个由个
37、由5个核苷酸组成的保守序列,个核苷酸组成的保守序列,是聚合酶结合位点,称为是聚合酶结合位点,称为Pribnow区区,其中央大约位于起点上游,其中央大约位于起点上游10bp处,所以又称为处,所以又称为10区区。-10序列序列 (Pribnow框盒框盒) 其保守序列为其保守序列为TATAAT,位于,位于-10bp左右,其左右,其中中3端的端的“T”十分保守。十分保守。 A.T较丰富,易于解链。较丰富,易于解链。 它和转录起始位点一般相距它和转录起始位点一般相距5bp。 功能功能: RNA pol紧密结合紧密结合; 形成开放启动复合体;形成开放启动复合体;(1) 使使RNA pol定向转录。定向转录
38、。如果把如果把Pribnow区从区从TATAAT变成变成AATAAT就会使就会使该启动子发生该启动子发生下降突变下降突变(down mutation);如果增加如果增加Pribnow区的共同序列,将乳糖操纵子的区的共同序列,将乳糖操纵子的启动子中的启动子中的TATGTT变成变成TATATT,就会提高启动,就会提高启动子的效率,称为子的效率,称为上升突变上升突变(up mutation)。提纯被保护的片段后却提纯被保护的片段后却发现,发现,RNA聚合酶并不聚合酶并不能重新结合或并不能选能重新结合或并不能选择正确的起始点,表明择正确的起始点,表明在保护区外可能还存在在保护区外可能还存在与与RNA聚
39、合酶对启动子聚合酶对启动子的识别有关的序列。的识别有关的序列。 ? 科学家又从噬菌体的左、右启动子科学家又从噬菌体的左、右启动子PL及及PR和和SV40启动子的启动子的35 bp附近找到了附近找到了另一段共同序列:另一段共同序列:TTGACA。35区的发现区的发现-35序列序列 (Sextama盒盒) 其保守序列为其保守序列为TTGACA, 与与-10序列相隔序列相隔16-19bp。 功能功能: (1)为为RNA pol的识别位点。的识别位点。 (2)RNA Pol的核心酶只能起到与模板结合和的核心酶只能起到与模板结合和催化的功能,并不能识别催化的功能,并不能识别-35序列,只有序列,只有亚亚
40、基才能识别基才能识别-35序列,为转录选择模板链。序列,为转录选择模板链。 35区区 10区区T85T83G81A61C69A52T89A89T50A65A100 大部分启动子都存在这两段共同序列,即位大部分启动子都存在这两段共同序列,即位于于10 bp处的处的TATA区和区和35 bp处的处的TTGACA区。区。 它们是它们是RNA聚合酶与启动子的结合位点聚合酶与启动子的结合位点,能与能与因子相互识别而具有很高的亲和力。因子相互识别而具有很高的亲和力。 100个个E.coli的不同启动子的序列测定结果的不同启动子的序列测定结果10区和区和35区的最佳距离区的最佳距离 在原核生物中在原核生物中
41、, 10区和区和35区的最佳距离区的最佳距离大约是大约是1619 bp。 过大或过小都会降低转录活性。过大或过小都会降低转录活性。 这可能是因为这可能是因为RNA Pol本身的大小和空间结本身的大小和空间结构有关。构有关。geneTTGACAAACTGTTATAATPuATATTAPytranscribed initiation site -50 -40 -30 -20 -10 1 10 5Recognition siteBinding sitePribnow boxpromoter 40-80bp123(1)结构典型,都含有识别)结构典型,都含有识别(R),结合,结合(B)和起始和起始(I)
42、三个位点;三个位点;(2)序列保守,如)序列保守,如-35序列,序列,-10序列结构都十分保守;序列结构都十分保守;(3)位置和距离都比较恒定;)位置和距离都比较恒定;(4)直接和多聚酶相结合;)直接和多聚酶相结合;(5)常和操纵子相邻;)常和操纵子相邻;(6)都在其控制基因的)都在其控制基因的5端;端;(7)决定转录的启动和方向。)决定转录的启动和方向。原核生物启动子原核生物启动子的共同的特点的共同的特点真核生物启动子的结构特点真核生物启动子的结构特点真核生物真核生物RNA聚合酶聚合酶II所识别的启动子区所识别的启动子区 Hogness等发现类似等发现类似Pribnow区的区的Hogness
43、区区,在转录起始点上游在转录起始点上游2530 bp处,保守序列处,保守序列为为TATAAA,也称,也称TATA区区。在起始位点上游在起始位点上游7078 bp处还有另一段共同处还有另一段共同序列序列CCAAT,称为,称为CAAT区区(CAAT box)。)。1. 核心元件核心元件TATA box: 2530 bp区区启始子启始子(initiator,Inr) :转录起始位点附近:转录起始位点附近真核基因的启动子真核基因的启动子2. 上游启动子元件(上游启动子元件(UPE) CAAT box :7080区区CCAAT序列序列GC box: 80110含有含有GCCACACCC 或或GGGCGG
44、G序列。序列。 上游启动子元件上游启动子元件(upstream promoter element,UPE):): 将将TATA区上游的保守序列称为上游启动子区上游的保守序列称为上游启动子元件或称元件或称上游激活序列上游激活序列(upstream activating sequence,UAS)。)。 TATA区区-使转录精确地起始使转录精确地起始:如果除去如果除去TATA区或进行碱基突变,转录产物下降区或进行碱基突变,转录产物下降的相对值不如的相对值不如CAAT区或区或GC区突变后明显,但发区突变后明显,但发现所获得的现所获得的RNA产物起始点不固定。产物起始点不固定。真核生物启动子对转录的影
45、响真核生物启动子对转录的影响 CAAT区和区和GC区主要控制转录起区主要控制转录起始频率:始频率:基本不参与起始位点的确定。基本不参与起始位点的确定。研究研究SV40晚期基因启动子发现,上游激活区的存在与否,对该晚期基因启动子发现,上游激活区的存在与否,对该启动子的生物活性有着根本性的影响。若将该基因启动子的生物活性有着根本性的影响。若将该基因5 上游上游2147核苷酸序列切除,基因完全不表达。核苷酸序列切除,基因完全不表达。增强子增强子(enhancer)1981年由年由Benerji, Rusconi小组和小组和Chambom等发现的,又称等发现的,又称远上游序列远上游序列(far ups
46、tream sequence)。)。已在已在SV40的转录单元上发现其转录起始位点的转录单元上发现其转录起始位点上游约上游约200bp处有两段处有两段72bp长的重复序列,长的重复序列,它们不是启动子的一部分,但能增强或促进它们不是启动子的一部分,但能增强或促进转录的起始,因此,称这种能强化转录起始转录的起始,因此,称这种能强化转录起始的序列为的序列为增强子增强子或或强化子强化子(enhancer)。)。远端调控区远端调控区 增强子(增强子(Enhancer )增强子很可能通过影响染色质增强子很可能通过影响染色质DNA蛋白质结构或改变蛋白质结构或改变超螺旋的密度而改变模板的整体结构,从而使得超
47、螺旋的密度而改变模板的整体结构,从而使得RNA聚聚合酶更容易与模板合酶更容易与模板DNA相结合,起始基因转录。相结合,起始基因转录。增强子无论位于靶基因的上游、下游增强子无论位于靶基因的上游、下游或是内部都可发挥增强转录的作用。或是内部都可发挥增强转录的作用。增强子特点增强子特点 具有具有远距离效应远距离效应。 常在上游常在上游-200bp处,但可增强远处启动子的转录,即使相距十几处,但可增强远处启动子的转录,即使相距十几Kb也能发也能发挥其作用;挥其作用; 无方向性。无方向性。 无论在靶基因的上游,下游或内部都可发挥增强转录的作用;无论在靶基因的上游,下游或内部都可发挥增强转录的作用; 顺式
48、调节顺式调节。 只调节位于同一染色体上的靶基因,而对其它染色体上的基因无作用;只调节位于同一染色体上的靶基因,而对其它染色体上的基因无作用; 无物种和基因的特异性无物种和基因的特异性, 可以接到异源基因上发挥作用可以接到异源基因上发挥作用; 具有具有组织的特异性组织的特异性。 SV40的增强子在的增强子在3T3细胞中比多瘤病毒的增强子要弱,但在细胞中比多瘤病毒的增强子要弱,但在HeLa细胞中细胞中SV40的增强子比多瘤病毒的要强的增强子比多瘤病毒的要强5倍。增强子的效应需特定的蛋白质因子倍。增强子的效应需特定的蛋白质因子参与。参与。 有有相位性相位性。 其作用和其作用和DNA的构象有关。的构象
49、有关。 有的增强子可以有的增强子可以对外部信号产生反应对外部信号产生反应。 如热休克基因在高温下才表达。编码重金属蛋白的金属硫蛋白基因在镉和如热休克基因在高温下才表达。编码重金属蛋白的金属硫蛋白基因在镉和锌存在下才表达。某些增强子可以被固醇类激素所激活。锌存在下才表达。某些增强子可以被固醇类激素所激活。 真核生物的启动子特点真核生物的启动子特点 (1)有多种元件:)有多种元件:TATA框,框,GC框,框,CAAT框等;框等;(2)结构不恒定。如)结构不恒定。如SV40早期基因早期基因,缺少缺少TATA和和CAAT,但有,但有6个个GC区区;组蛋白组蛋白H2B,不含不含GC区区,但有两个但有两个
50、CAAT区一个区一个TATA区区;(3)它们的位置、序列、距离和方向都不完全相同)它们的位置、序列、距离和方向都不完全相同;(4)有的有远距离的调控元件存在,如增强子;)有的有远距离的调控元件存在,如增强子;(5)这些元件常常起到控制转录效率和选择起始位点的作用;)这些元件常常起到控制转录效率和选择起始位点的作用;(6)不直接和)不直接和RNA pol结合。转录时先和其它转录激活因子相结结合。转录时先和其它转录激活因子相结合,再和聚合酶结合。合,再和聚合酶结合。TATA boxHogness boxpromoter gene-50 -40 -30 -20 -10 1 10 5CAAT boxG
51、C box原核与真核生物原核与真核生物mRNA的特征比较的特征比较原核生物原核生物中中: mRNA的转录和翻译发生在的转录和翻译发生在同一个细胞空间同一个细胞空间, 这两个过程几乎是这两个过程几乎是同步同步进行的进行的 。真核细胞中:真核细胞中:真核细胞真核细胞mRNA的合成和功能表达发生在不同的合成和功能表达发生在不同的空间和时间范畴内。的空间和时间范畴内。 mRNA以较大分子量的以较大分子量的前体前体RNA出现在核内,出现在核内,只有成熟的、相对分子质量明显变小并经化学只有成熟的、相对分子质量明显变小并经化学修饰的修饰的mRNA才能进入细胞质,参与蛋白质的才能进入细胞质,参与蛋白质的合成。
52、合成。mRNA的组成:的组成:编码区编码区(coding region):从起始密码子从起始密码子AUG开开始经一连串编码氨基酸的密码子直至终止密码始经一连串编码氨基酸的密码子直至终止密码子。子。5端上游非编码区端上游非编码区(5UTR):位于位于AUG之前不之前不翻译的区域。翻译的区域。3端下游非编码区端下游非编码区(3UTR) :位于终止密码子位于终止密码子之后不翻译的区域。之后不翻译的区域。原核生物原核生物mRNA的特征的特征 半衰期短。半衰期短。 许多原核生物许多原核生物mRNA以多顺反子的形式以多顺反子的形式存在。存在。 原核生物原核生物mRNA的的5端无帽子结构,端无帽子结构,3端
53、端没有或只有较短的多聚(没有或只有较短的多聚(A)结构。)结构。 半衰期短半衰期短原核生物原核生物mRNA的特征的特征原核生物中,原核生物中,mRNA的的转录和翻译是在同一个转录和翻译是在同一个细胞空间里同步进行细胞空间里同步进行的,的,蛋白质合成往往在蛋白质合成往往在mRNA刚开始转录时就刚开始转录时就被引发了。被引发了。大多数细菌大多数细菌mRNA在转在转录开始录开始1分钟后就开始降分钟后就开始降解。解。mRNA降解的速度大降解的速度大概只有转录或翻译速度的概只有转录或翻译速度的一半。一半。2. 许多以许多以多顺反子多顺反子的形式存在:的形式存在:原核细胞的原核细胞的mRNA(包括病毒包括
54、病毒)有时可以同有时可以同时编码几个多肽。时编码几个多肽。原核生物原核生物mRNA的特征的特征Prokaryotic mRNA (polycistrionic)单顺反子单顺反子mRNA (monocistronic mRNA): 只编码一个蛋白质的只编码一个蛋白质的mRNA。多顺反子多顺反子mRNA(polycistronic mRNA):编码多个蛋白质的编码多个蛋白质的mRNA。 3. 原核生物原核生物mRNA的的5端无帽子结构,端无帽子结构,3端没有或只有较短的多聚(端没有或只有较短的多聚(A)结构。)结构。原核生物原核生物mRNA的特征的特征原核生物起始密码子原核生物起始密码子AUG上游
55、有一被称为上游有一被称为Ribosome Binding Site (RBS)或或SD序列序列(Shine Dalgarno sequence)的保守区,因为该)的保守区,因为该序列序列与与16S-rRNA 3端反向互补端反向互补,所以被认为在核糖体,所以被认为在核糖体-mRNA的的结合过程中起作用。结合过程中起作用。 4. 原核生物原核生物常以常以AUG(有时(有时GUG,甚至,甚至UUG)作为)作为起始密码子起始密码子; 真核生物几乎永远以真核生物几乎永远以AUG作为起始密作为起始密 码子。码子。原核生物原核生物mRNA的特征的特征 单顺反子形式存在。单顺反子形式存在。 5端端存在存在“帽
56、子帽子”结构。结构。 绝大多数具有绝大多数具有多聚多聚(A)尾巴尾巴。真核生物真核生物mRNA的特征的特征真核生物真核生物mRNA的结构模式的结构模式Eukaryotic mRNA (monocistrionic)“基因基因”的分子生物学定义是:产的分子生物学定义是:产生一条多肽链或功能生一条多肽链或功能RNARNA所必需的所必需的全全部部核苷酸序列!核苷酸序列!A gene can be defined as following: The entire nucleic acid sequence that is necessary for the synthesis of a functio
57、nal polypeptide or RNA molecule. 真核生物真核生物mRNA的的5端端存在存在“帽帽子子”结构。结构。 真核生物基因转录一般从嘌呤起始,真核生物基因转录一般从嘌呤起始,其其5端大都经过修饰。端大都经过修饰。真核生物真核生物mRNA的特征的特征5Capping 通过通过 5 5磷酸二酯键在原初磷酸二酯键在原初mRNA的的5端倒端倒扣一个扣一个“G”。 在新生在新生mRNA链达到链达到50个核苷酸前,甚至可个核苷酸前,甚至可能在能在RNA Pol II 离开转录起始位点之前,帽子离开转录起始位点之前,帽子结构就已加到结构就已加到mRNA的第一个核苷酸上了。的第一个核苷
58、酸上了。 5末端加上鸟苷是由末端加上鸟苷是由鸟苷转移酶催化鸟苷转移酶催化的的 。 帽子结构是帽子结构是GTP和原和原5三磷酸腺苷(或鸟苷)三磷酸腺苷(或鸟苷)缩合反应的产物。缩合反应的产物。 mRNA的帽子结构常常的帽子结构常常被甲基化被甲基化。真核生物真核生物mRNA的的“帽子帽子”结构结构鸟苷酸鸟苷酸-7甲基转移酶甲基转移酶2-O-甲基转移酶甲基转移酶 mRNA的帽子结构常常被甲基化的帽子结构常常被甲基化零类帽子零类帽子(cap0):第一个甲基出现在所有真核细胞的):第一个甲基出现在所有真核细胞的mRNA中(单细胞真核生物中(单细胞真核生物mRNA主要是这个结构),主要是这个结构),由由鸟
59、苷酸鸟苷酸-7甲基转移酶甲基转移酶催化,称为零类帽子。催化,称为零类帽子。 1类帽子类帽子(cap1):如在:如在第二个核苷酸第二个核苷酸(原原mRNA 5第一位第一位)的的2-OH位上加另一个甲基位上加另一个甲基,这步反应由,这步反应由2-O-甲基转移甲基转移酶酶完成。一般把有这两个甲基的结构称为完成。一般把有这两个甲基的结构称为1类帽子。类帽子。真核真核生物中以这类帽子结构为主。生物中以这类帽子结构为主。2类帽子类帽子(cap2): 在某些生物细胞内,在某些生物细胞内,mRNA链上的链上的第三第三个核苷酸的个核苷酸的2-OH位也可能被甲基化位也可能被甲基化,因为这个反应只,因为这个反应只以
60、带有以带有1类帽子的类帽子的mRNA为底物,所以被称为为底物,所以被称为2类帽子。类帽子。只占有帽只占有帽mRNA总量的总量的10%-15%以下。以下。帽子结构的功能帽子结构的功能(1)有助于有助于mRNA越过核膜,进入胞质;越过核膜,进入胞质;(2)保护保护5不被核酶降解;不被核酶降解;(3)翻译时供翻译时供IF(起始因子)和核糖体识(起始因子)和核糖体识别,是翻译所必需的。别,是翻译所必需的。 2. 绝大多数绝大多数真核生物真核生物mRNA具有具有多聚多聚(A)尾巴尾巴。除组蛋白基因外,真核生物除组蛋白基因外,真核生物mRNA的的3末末端端都有多聚都有多聚(A)序列,其长度因序列,其长度因mRNA种类不同
温馨提示
- 1. 本站所有资源如无特殊说明,都需要本地电脑安装OFFICE2007和PDF阅读器。图纸软件为CAD,CAXA,PROE,UG,SolidWorks等.压缩文件请下载最新的WinRAR软件解压。
- 2. 本站的文档不包含任何第三方提供的附件图纸等,如果需要附件,请联系上传者。文件的所有权益归上传用户所有。
- 3. 本站RAR压缩包中若带图纸,网页内容里面会有图纸预览,若没有图纸预览就没有图纸。
- 4. 未经权益所有人同意不得将文件中的内容挪作商业或盈利用途。
- 5. 人人文库网仅提供信息存储空间,仅对用户上传内容的表现方式做保护处理,对用户上传分享的文档内容本身不做任何修改或编辑,并不能对任何下载内容负责。
- 6. 下载文件中如有侵权或不适当内容,请与我们联系,我们立即纠正。
- 7. 本站不保证下载资源的准确性、安全性和完整性, 同时也不承担用户因使用这些下载资源对自己和他人造成任何形式的伤害或损失。
最新文档
- 泰山护理职业学院《计算机电路基础》2023-2024学年第二学期期末试卷
- 张家口职业技术学院《接口自动化》2023-2024学年第二学期期末试卷
- 贵州铜仁数据职业学院《桥梁结构非线性》2023-2024学年第一学期期末试卷
- 山东英才学院《儿童文学(小教)》2023-2024学年第二学期期末试卷
- 郑州经贸学院《钢琴弹唱》2023-2024学年第一学期期末试卷
- 湖南师范大学《公共健康与预防医学》2023-2024学年第二学期期末试卷
- 反担保保证抵押借款合同
- 抵押物品的合同
- 手房买卖合同独家合同
- 畜牧产品产销对接与供应链保障合同
- 前列腺增生患者的护理查房课件
- 2023年四川农信(农商行)招聘笔试真题
- 呼兰河传(2022年黑龙江牡丹江中考语文试卷记叙文阅读题及答案)
- 小学英语“教学评一体化”实施
- 危险化学品出入库管理制度
- 人教版道德与法治三年级下册全册课件(完整版)
- 医学课件新生儿黄疸5
- 2024年中考英语作文热点主题:人工智能满分范文10篇精彩表达25句
- 2025年呼和浩特市重点中学中考领航2020大二轮复习数学试题模拟含解析
- 水泥搅拌桩施工记录-自动计算
- 全国大学生网络安全知识竞赛题库及答案
评论
0/150
提交评论